ERN1 eli IRE1beta
ASK1
https://pmc.ncbi.nlm.nih.gov/articles/PMC9038009/
Kesken

PHYTIN, Fytiini (IP6) An interesting molecule in human food and the human biochemistry.
https://pubmed.ncbi.nlm.nih.gov/35112133/
PubMed HAKU: Na+, K+, Cl-, ENDOLYMPHA.
Abstract
The endolymphatic sac is a small sac-shaped organ at the end of the membranous labyrinth of the inner ear. The endolymphatic sac absorbs the endolymph, in which the ion balance is crucial for inner ear homeostasis. Of the three sections of the endolymphatic sac, the intermediate portion is the center of endolymph absorption, particularly sodium transport, and is thought to be regulated by aldosterone. Disorders of the endolymphatic sac may cause an excess of endolymph (endolymphatic hydrops), a histological observation in Meniere’s disease. A low-salt diet is an effective treatment for Meniere’s disease, and is based on the assumption that the absorption of endolymph in the endolymphatic sac abates endolymphatic hydrops through a physiological increase in aldosterone level. However, the molecular basis of endolymph absorption in each portion of the endolymphatic sac is largely unknown because of difficulties in gene expression analysis, resulting from its small size and intricate structure. The present study combined reverse transcription-quantitative polymerase chain reaction and laser capture microdissection techniques to analyze the difference of gene expression of the aldosterone-controlled epithelial Na+ channel, thiazide-sensitive Na+-Cl− cotransporter, and Na+, K+-ATPase genes in the three individual portions of the endolymphatic sac in a rat model. A low-salt diet increased the expression of aldosterone-controlled ion transporters, particularly in the intermediate portion of the endolymphatic sac. Our findings will contribute to the understanding of the physiological function of the endolymphatic sac and the pathophysiology of Meniere’s disease.
The cochlear stria vascularis produces endolymph and thereby plays an active role in inner ear homeostasis. We recently reported that the H+/myo-inositol cotransporter (HMIT) gene is expressed in the stria vascularis. Here, we examined the protein localization of HMIT and Na+/myo-inositol cotransporter 1 (SMIT1) in the stria vascularis by immunohistochemistry. HMIT and SMIT1 were detected in the lateral wall of the cochlear duct. HMIT was widely detected throughout the stria vascularis, while SMIT1 was enriched in the strial basal cells. To examine the localization of HMIT in the stria vascularis in more detail, dissociated strial cells were immunostained, which resulted in the detection of HMIT immunoreactivity in marginal cells. These results indicate that HMIT is expressed in marginal cells and basal cells of the stria vascularis, while SMIT1 expression is enriched in basal cells. We speculate that HMIT and SMIT1 may play important roles in the homeostasis of cochlear fluids, for example by participating in pH regulation and osmoregulation.
Keywords: Endolymph; Inner ear; Osmoregulation; Rat (Brown Norway); pH.
Genes: GeneCards
HMIT, Alias SLC2A13 (12q12)
https://www.genecards.org/cgi-bin/carddisp.pl?gene=SLC2A13&keywords=HMIT
Enables ATPase binding activity; myo-inositol:proton symporter activity;
and protease binding activity. Involved in myo-inositol transport and
positive regulation of amyloid-beta formation. Is integral component of
plasma membrane. Part of cell body; cell periphery; and cell projection.
[provided by Alliance of Genome Resources, Apr 2022]
SMIT1 (21q22.11)
https://www.genecards.org/Search/Keyword?queryString=SMIT1
Enables potassium channel regulator activity and transmembrane transporter binding activity. Predicted to be involved in inositol metabolic process; monosaccharide transmembrane transport; and myo-inositol import across plasma membrane. Predicted to act upstream of or within several processes, including peripheral nervous system development; positive regulation of reactive oxygen species biosynthetic process; and regulation of respiratory gaseous exchange. Located in plasma membrane. Part of perinuclear region of cytoplasm. [provided by Alliance of Genome Resources, Apr 2022]
Electrogenic Na(+)-coupled sugar symporter that actively transports myo-inositol and its stereoisomer scyllo-inositol across the plasma membrane, with a Na(+) to sugar coupling ratio of 2:1 (By similarity). Maintains myo-inositol concentration gradient that defines cell volume and fluid balance during osmotic stress, in particular in the fetoplacental unit and central nervous system (By similarity). Forms coregulatory complexes with voltage-gated K(+) ion channels, allosterically altering ion selectivity, voltage dependence and gating kinetics of the channel. In turn, K(+) efflux through the channel forms a local electrical gradient that modulates electrogenic Na(+)-coupled myo-inositol influx through the transporter (PubMed:24595108, 28793216). Associates with KCNQ1-KCNE2 channel in the apical membrane of choroid plexus epithelium and regulates the myo-inositol gradient between blood and cerebrospinal fluid with an impact on neuron excitability (By similarity) (PubMed:24595108). Associates with KCNQ2-KCNQ3 channel altering ion selectivity, increasing Na(+) and Cs(+) permeation relative to K(+) permeation (PubMed:28793216). Provides myo-inositol precursor for biosynthesis of phosphoinositides such as PI(4,5)P2, thus indirectly affecting the activity of phosphoinositide-dependent ion channels and Ca(2+) signaling upon osmotic stress (PubMed:27217553). ( SC5A3_HUMAN,P53794 )
KCNQ1-KCNE2 channel KCNQ1-KCNE2 channel associates with Na(+)-coupled myo-inositol symporter in the apical membrane of choroid plexus epithelium and regulates the myo-inositol gradient between blood and cerebrospinal fluid with an impact on neuron excitability (By similarity). ( KCNE2_HUMAN,Q9Y6J6 )
Potassium channel that plays an important role in a number of tissues, including heart, inner ear, stomach and colon (PubMed:10646604, 25441029). Associates with KCNE beta subunits that modulates current kinetics (PubMed:10646604, 11101505, 19687231, 8900283, 9108097, 9312006). Induces a voltage-dependent current by rapidly activating and slowly deactivating potassium-selective outward current (PubMed:10646604, 11101505, 25441029, 8900283, 9108097, 9312006). Promotes also a delayed voltage activated potassium current showing outward rectification characteristic (By similarity). During beta-adrenergic receptor stimulation participates in cardiac repolarization by associating with KCNE1 to form the I(Ks) cardiac potassium current that increases the amplitude and slows down the activation kinetics of outward potassium current I(Ks) (By similarity) (PubMed:10646604, 11101505, 8900283, 9108097, 9312006). Muscarinic agonist oxotremorine-M strongly suppresses KCNQ1/KCNE1 current (PubMed:10713961). When associated with KCNE3, forms the potassium channel that is important for cyclic AMP-stimulated intestinal secretion of chloride ions (PubMed:10646604). This interaction with KCNE3 is reduced by 17beta-estradiol, resulting in the reduction of currents (By similarity). During conditions of increased substrate load, maintains the driving force for proximal tubular and intestinal sodium ions absorption, gastric acid secretion, and cAMP-induced jejunal chloride ions secretion (By similarity). Allows the provision of potassium ions to the luminal membrane of the secretory canaliculus in the resting state as well as during stimulated acid secretion (By similarity). When associated with KCNE2, forms a heterooligomer complex leading to currents with an apparently instantaneous activation, a rapid deactivation process and a linear current-voltage relationship and decreases the amplitude of the outward current (PubMed:11101505). When associated with KCNE4, inhibits voltage-gated potassium channel activity (PubMed:19687231). When associated with KCNE5, this complex only conducts current upon strong and continued depolarization (PubMed:12324418). Also forms a heterotetramer with KCNQ5; has a voltage-gated potassium channel activity (PubMed:24855057). Binds with phosphatidylinositol 4,5-bisphosphate (PubMed:25037568). KCNQ1-KCNE2 channel associates with Na(+)-coupled myo-inositol symporter in the apical membrane of choroid plexus epithelium and regulates the myo-inositol gradient between blood and cerebrospinal fluid with an impact on neuron excitability. ( KCNQ1_HUMAN,P51787 )
Interacts with KCNE2; forms a heterooligomer complex that targets to the membrane raft and leading to currents with an apparently instantaneous activation, a rapid deactivation process and a linear current-voltage relationship and decreases the amplitude of the outward current (PubMed:11101505, 20533308). -> KCNE2 (Gene 21q22.11)
HAIR CELLS
https://pubmed.ncbi.nlm.nih.gov/34566562/
JONITASAPAINOSTA ENDOLYM;FASSA
ENDOLYMFASTA : Sisäkorvan miljööstä.
Abstract
Under normal conditions, the inner ear possesses remarkably stable homeostatic mechanisms for the maintenance of functional integrity of the inner ear fluid. The inner ear fluid maintains its homeostasis by a variety of regulatory mechanisms such as an ion transport system, a blood-labyrinth barrier, and a constant blood supply. Highly regulated transport of ions into and out of the inner ear provides for the maintenance of inner ear fluid composition necessary for auditory transduction. Any disturbance in one of these mechanisms can disrupt homeostasis expressed by ionic, osmotic, or metabolic imbalance between the compartments. Free radicals, stress hormones, noise exposure, and aminoglycoside antibiotics may induce short- and long-term effects on cellular function of the auditory or vestibular system (or both) and serve as a triggering mechanism for abrupt functional disturbances of inner ear fluid ion homeostasis. In this article, we present a comprehensive review of the mechanisms underlying inner ear fluid homeostasis necessary for normal auditory function and factors that can disrupt homeostasis and lead to functional disturbances, namely sensorineural hearing loss, tinnitus, and vertigo.
MIKÄ ON INOSITOLIN OSUUS ENDOLYMFAMILJÖÖN HOMEOSTAASISSA? Katso seuraava artikkeli.Endolymfa ja osmoottinen stressi. Inositolilla funktiota .
Phosphatidylinositol 3-kinase is composed of an 85 kDa regulatory subunit and a 110 kDa catalytic subunit. The protein encoded by this gene represents the catalytic subunit, which uses ATP to phosphorylate PtdIns, PtdIns4P and PtdIns(4,5)P2. This gene has been found to be oncogenic and has been implicated in cervical cancers. A pseudogene of this gene has been defined on chromosome 22. [provided by RefSeq, Apr 2016]
PIK3CA is the most recurrently mutated gene in breast cancer, and has been found to important in a number of cancer types. An integral part of the PI3K pathway, PIK3CA has long been described as an oncogene, with two main hotspots for activating mutations, the 542/545 region of the helical domain, and the 1047 region of the kinase domain. PIK3CA, and its interaction with the AKT and mTOR pathways, is the subject of an immense amount of research and development, and PI3K inhibition has seen some limited success in recent clinical trials. While monotherapies seem to be limited in their potential, there is a recent interest in pursuing PI3K inhibition as part of a combination therapy regiment with inhibition partners including TKI's, MEK inhibitors, PARP inhibitors, and in breast cancer, aromatase inhibitors.
PIK3CA (Phosphatidylinositol-4,5-Bisphosphate 3-Kinase Catalytic Subunit Alpha) is a Protein Coding gene. Diseases associated with PIK3CA include Megalencephaly-Capillary Malformation-Polymicrogyria Syndrome and Congenital Lipomatous Overgrowth, Vascular Malformations, And Epidermal Nevi. Among its related pathways are Downstream signaling of activated FGFR2 and Translation Insulin regulation of translation. Gene Ontology (GO) annotations related to this gene include transferase activity, transferring phosphorus-containing groups and protein serine/threonine kinase activity. An important paralog of this gene is PIK3CB.
Phosphoinositide-3-kinase (PI3K) phosphorylates phosphatidylinositol (PI) and its phosphorylated derivatives at position 3 of the inositol ring to produce 3-phosphoinositides (PubMed:15135396, 23936502, 28676499). Uses ATP and PtdIns(4,5)P2 (phosphatidylinositol 4,5-bisphosphate) to generate phosphatidylinositol 3,4,5-trisphosphate (PIP3) (PubMed:15135396, 28676499). PIP3 plays a key role by recruiting PH domain-containing proteins to the membrane, including AKT1 and PDPK1, activating signaling cascades involved in cell growth, survival, proliferation, motility and morphology. Participates in cellular signaling in response to various growth factors. Involved in the activation of AKT1 upon stimulation by receptor tyrosine kinases ligands such as EGF, insulin, IGF1, VEGFA and PDGF. Involved in signaling via insulin-receptor substrate (IRS) proteins. Essential in endothelial cell migration during vascular development through VEGFA signaling, possibly by regulating RhoA activity. Required for lymphatic vasculature development, possibly by binding to RAS and by activation by EGF and FGF2, but not by PDGF. Regulates invadopodia formation through the PDPK1-AKT1 pathway. Participates in cardiomyogenesis in embryonic stem cells through a AKT1 pathway. Participates in vasculogenesis in embryonic stem cells through PDK1 and protein kinase C pathway. In addition to its lipid kinase activity, it displays a serine-protein kinase activity that results in the autophosphorylation of the p85alpha regulatory subunit as well as phosphorylation of other proteins such as 4EBP1, H-Ras, the IL-3 beta c receptor and possibly others (PubMed:23936502, 28676499). Plays a role in the positive regulation of phagocytosis and pinocytosis (By similarity). ( PK3CA_HUMAN,P42336 )
PI 3-Kinases (phosphoinositide 3-kinases, PI 3-Ks) are a family of lipid kinases capable of phosphorylating the 3'OH of the inositol ring of phosphoinositides. They are responsible for coordinating a diverse range of cell functions including proliferation and survival.
2020
https://www.nature.com/articles/s41586-020-2969-2
Nature volume 588, pages 331–336 (2020) Abstract
Most deaths from cancer are explained by metastasis, and yet large-scale metastasis research has been impractical owing to the complexity of in vivo models. Here we introduce an in vivo barcoding strategy that is capable of determining the metastatic potential of human cancer cell lines in mouse xenografts at scale. We validated the robustness, scalability and reproducibility of the method and applied it to 500 cell lines1,2 spanning 21 types of solid tumour. We created a first-generation metastasis map (MetMap) that reveals organ-specific patterns of metastasis, enabling these patterns to be associated with clinical and genomic features. We demonstrate the utility of MetMap by investigating the molecular basis of breast cancers capable of metastasizing to the brain—a principal cause of death in patients with this type of cancer. Breast cancers capable of metastasizing to the brain showed evidence of altered lipid metabolism. Perturbation of lipid metabolism in these cells curbed brain metastasis development, suggesting a therapeutic strategy to combat the disease and demonstrating the utility of MetMap as a resource to support metastasis research..
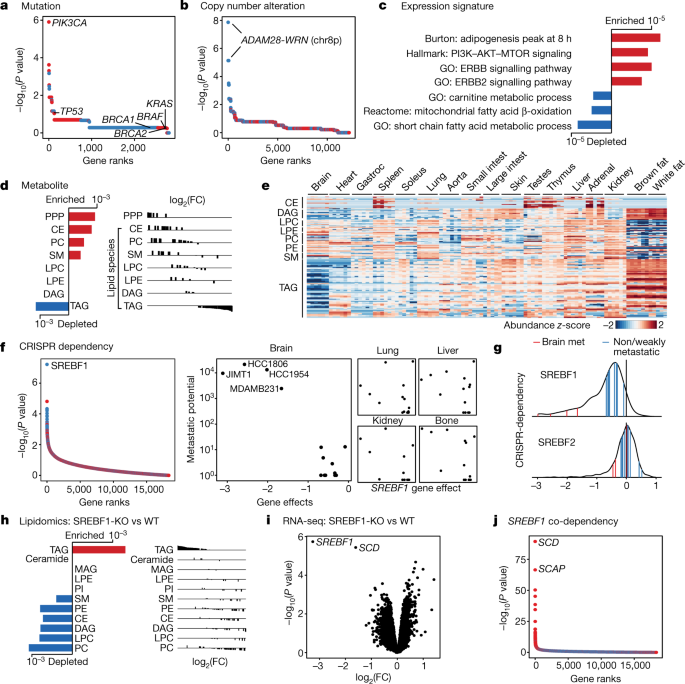
a, Somatic mutations that associate with brain metastatic potential in the basal-like breast cancer cohort. The top correlate, PIK3CA, reaches statistical significance (FDR = 0.0034, highlighted in bold). All PIK3CA mutations are activating. Positive correlations are in red, negative correlations are in blue. Selected known oncogenes or tumour suppressors in basal-like breast cancer are presented for comparison. b, Alterations in copy number that associate with brain metastatic potential. The top correlates cluster in chr 8p12–8p21.2 (FDR = 0.0017, highlighted in bold). c, Gene-expression signatures that associate with brain metastatic potential. Bars indicate P values. Expression signature scores were projected for each cell line with their in vitro RNA-seq data and used for regression analysis. GO (Gene Ontology), Hallmark, Reactome and Burton are gene sets in the MSigDB gene set enrichment analysis (GSEA) collection. d, Lipid-metabolite species that associate with brain metastatic potential. Bars indicate P values. Lipid metabolites measured by mass spectrometry were grouped by species, and enrichment analysis of the species was performed using GSEA. CE, cholesterol ester; PC, phosphatidylcholine; SM, sphingomyelin; LPC, lysophosphatidylcholine; LPE, lysophosphatidylethanolamine; DAG, diacylglycerol; PPP, pentose phosphate pathway metabolites. e, Heat map presenting distribution of lipid species measured by mass spectrometry from different mouse tissues. Gastroc, gastrocnemius. f, CRISPR gene dependencies that associate with brain metastatic potential. The top gene, SREBF1 (FDR = 0.001), is a selective dependency in highly brain metastatic lines. Positive correlations are in red, negative correlations are in blue. g, Distribution of SREBF1 (top) and SREBF2 (bottom) dependencies across 688 human cancer cell lines. The positions of highly brain metastatic (met) breast lines are highlighted in red, whereas weakly metastatic or non-brain metastatic breast lines are highlighted in blue. h, Consensus alterations in lipid species abundance upon SREBF1 knockout (KO) in JIMT1 and HCC1806, two brain metastatic cell lines. Bars indicate adjusted P values. Lipid metabolites measured by mass spectrometry were grouped by species, and enrichment analysis of the species was performed using GSEA. WT, wild type. i, Consensus gene-expression changes upon SREBF1 knockout in JIMT1, HCC1806, HCC1954 and MDAMB231, four brain metastatic cell lines. The two top genes are SREBF1 and SCD (FDR <0.05, highlighted in bold). j, Co-dependencies of SREBF1 across 688 human cancer cell lines in genome-wide CRISPR viability screen. The two top genes are SCD and SCAP (FDR < 1 × 10−60, highlighted in bold).
Given the observation that SREBF1 knockout resulted in a viability defect in vitro (Extended Data Fig. 10a), we compared the relative effect of knockout on metastasis to different organs, to determine whether the viability defect was preferentially observed in brain (Fig. 5d). Five weeks following intracardiac injection of SREBF1-knockout cells, we observed a marked defect in brain metastasis (196-fold reduction), compared with a modest defect in other organs (9–21 fold) (Fig. 5d). Histologic examination of brains from xenografted mice revealed large metastatic lesions in mice receiving wild-type cells, whereas those receiving SREBF1-knockout cells contained micrometastases (Extended Data Fig. 10b), suggesting that SREBF1 is not required for seeding the brain, but rather for proliferation in the brain microenvironment. Consistent with this hypothesis, injection of tumour cells into the carotid artery increased the probability of seeding the brain, but nevertheless a marked growth defect was still observed in SREBF1-knockout cells (Fig. 5e).
To determine the generality of the SREBF1 requirement for breast cancer growth in the brain, we knocked out SREBF1 in additional brain metastatic lines including HCC1954, MDAMB231 and HCC1806 using CRISPR–Cas9. As with JIMT1, a significant inhibition in brain metastatic growth was also observed in these lines, although the magnitude and duration of growth inhibition varied (Extended Data Fig. 10c, d). The least responsive cell line was HCC1806, in which SREBF1-knockout cells displayed a brain growth defect for the first week, but then assumed a growth trajectory that paralleled wild-type cells. This restoration of growth was not explained by reversion of the genome editing, as brain metastases at the end of the experiment showed evidence of editing at the SREBF1 locus and minimal SREBF1 protein expression (Extended Data Fig. 10e, f). Instead, we found that the SREBF1-independent growth was associated with upregulation of the fatty acid transporter CD36 and the fatty acid-binding protein FABP6 (Extended Data Fig. 10g). Of note, culture of HCC1806 in mouse brain-slice-conditioned medium similarly resulted in upregulation of SCD and CD36 expression (Extended Data Fig. 10h, i). JIMT1 cells did not upregulate CD36 or FABP6 expression following SREBF1 knockout (Extended Data Fig. 10g), perhaps explaining their inability to survive in the brain. Together, these results further demonstrate the relationship between lipid metabolism and brain metastasis, as cells under the selective pressure of SREBF1 loss must acquire lipids by other means to survive in the brain microenvironment.
This work describes MetMap as an approach for large-scale in vivo characterization of human cancer cell lines. The MetMap resource (available at https://pubs.broadinstitute.org/metmap) currently includes metastasis profiles of 500 cell lines spanning 21 tumour types, providing a large repertoire of models for exploration of metastasis mechanisms. A limitation of the use of human cell lines for such experiments is that they require the use of immunodeficient mice. The extent to which the immune system has a role in mediating patterns of metastasis remains to be determined37.
We followed up only a small proportion of the MetMap findings—specifically, breast cancer metastasis to brain. Multiple lines of experimental and clinical evidence pointed to a role of lipid metabolism in governing the ability of cells to survive in the brain microenvironment. The importance of lipid metabolism in cancer has been highlighted by a number of studies, but its role in brain metastasis has, to our knowledge, not been fully appreciated38,39,40,41. The possibility that interfering with lipid or cholesterol metabolism might abrogate metastatic growth in the brain is particularly intriguing. More generally, this work illustrates the complex interplay between cancer cell growth and the tissue microenvironment.
2023
https://www.nature.com/articles/s41467-023-44206-x
GSDensity can identify spatially relevant pathways in mouse brains and
human tumors including those following high-order organizational
patterns in the ST data. Particularly, we create a pan-cancer ST map
revealing spatially relevant and recurrently active pathways across six
different tumor types.
The proliferative ability of tumor cells could be regulated through their interactions with the TIME27,28. We then compared these actively dividing cells with other tumor cells from the angle of cell-cell interaction between tumor and TIME. We annotated the normal cells in the TNBC-1 dataset into two groups, fibroblasts, and immune cells (Supplementary Fig. 6b, c) and inferred the ligand-receptor interactions29 between immune and tumor cells and between tumor and tumor cells. We displayed all the ligand receptor-pairs in Supplementary Fig. 7a and found that the dividing cells showed a distinct profile of the GAS6-TYRO3 axis. All the other ligand-receptor pairs showed differential enrichments in either immune-tumor interaction or tumor-tumor interaction, while GAS6-TYRO3 was the only pair that showed differential enrichments in both groups. We then found that both the tumor cells and the immune cells could express the ligand, GAS6, while only the dividing tumor cells displayed high expression of the receptor, TYRO3 (Fig. 3h), which indicated the specific activation of the TYRO3 downstream signaling in those cells. For confirmation, we also fetched actively dividing tumor cells from another two TNBC datasets25, TNBC-2 and TNBC-5, and these cells also consistently showed high relevance to glycolysis, mTORC1 signaling, and mitotic spindle (Supplementary Fig. 6d–k, p-value < 2.2e−16 for all groups, Chi-squared test). The high expression of TYRO3 in actively dividing cells was also observed in the TNBC-5, confirming the previous finding (p-value = 3.18e−8, Wilcoxon test, Supplementary Fig. 6l–m). We then investigated this TYRO3 expression pattern in another published cohort with 8 TNBC patient samples30. TYRO3 were lowly detected in 7 of the samples (detected in 1–8% of tumor cells). In the only sample (GSM4909284_TN-MH0114-T2) with relative high expression of TYRO3 (detected in 24% of tumor cells), the actively dividing cells showed higher expression of TYRO3 than other tumor cells (p-value = 0.039, Wilcoxon test, Supplementary Fig. 6n). These results indicated that the overall expression level of TYRO3 in breast cancer cells is highly patient specific, while the high-TYRO3 expressing samples always had TYRO3 preferably express in a small group of actively dividing cells. The GAS6-TYRO3 axis has been associated with tumor cell proliferation, malignancy, and anti-PD1/PD-L1 resistance in previous studies31,32,33,34,35. Thus, through the integration of data and prior knowledge using GSDensity, we postulated a potential role TYRO3 in TNBC proliferation using only a few TNBC samples with very sparse single-cell gene expression profiles and generated a testable hypothesis for further studies.
..
We applied GSDensity to a ST mouse forebrain dataset generated by the 10X Visium technology. We first clustered the data spots based on the transcriptome (Fig. 5a) and observed that all the clusters were also spatially segregated on the spatial map (Fig. 5b). Thus, the pathways with cluster-wise specificity would naturally display spatial relevance in this data. However, it is unclear whether there are high order organization of pathway activities across multiple clusters, which would be undetectable in cluster-centric analysis. To address this question, we first identified 727 GO biological process terms with coordination in the dataset using GSDensity. For each term, we calculated its spatial relevance and specificity for each cluster. The spatial relevance is quantified by KL-divergence between the pathway weighted kernel density estimation (KDE) and the equally weighted KDE (Methods). The specificity of a pathway for a cluster is quantified by a specificity score based on Jensen-Shannon divergence, with larger values being more specific. We then plotted the 727 GO terms with their spatial relevance and maximum cluster-wise specificity (Fig. 5c). As expected, the spatial relevance showed largely positive correlation with cluster-wise specificity. Among the GO terms with high cluster-wise specificity (Fig. 5c, red) are dopamine receptor signaling for Cluster 1, amyloid beta formation for Cluster 5, ARP2/3 complex mediated actin nucleation for Cluster 8/11, and oligodendrocyte development for Cluster 2 (Fig. 5d–g). Interestingly, we also observed some GO terms with high spatial relevance and low cluster-wise specificity (Fig. 5c, blue). The cells highly relevant to these terms consisted of data spots from multiple clusters with higher-order spatial organization (Fig. 5h–k). For example, positive regulation of cellular response to insulin stimulus appeared highly active in the spots close to the caudal side (Fig. 5i), while fatty acid oxidation to the ventral side (Fig. 5j). It has been known that insulin receptors are expressed in hypothalamus and hippocampus43 which are both located close to the caudal side of this anterior section. Although GSDensity was designed to perform cluster-independent data analysis, we demonstrated that the pathway activity calculation by GSDensity can be easily integrated with cell information, such as cluster partition or spatial coordinates, when available.
..
Like the scenario of the mouse brain data, some pathways were both highly spatially relevant and highly cell-type specific, for example, protein localization to synapse was highly specific to inhibitory neurons and sensory perception of smell was highly specific to excitatory neurons, both of which occupied localizations with spatial relevance on the map (Supplementary Fig. 10c, d).
..