
Olin katsomassa juuri deubikitinaasien ryhmää- (DUB) ryhmää ja siiä mainittiin Sirtuiinit deasetylaasit, joihin muutamilla on interaktiovaikutusta.
Koska sirtuiinit taas ovat minulle aika vieras asia, olen useana päivänä katsonut niitä seitsemää sirtuiinia (SIRT1-7) ja mitä ne tekevät. Tässä joutuu jälleen aivan elementaarisiin molekyyleihin, sinne etikkahapon tasolle
ja koentsyymien ympyröihin.
Koentsyymeistä tietysti on hyvät rinkulat olemassa jo 1960-luvun oppikirjoissa siitä kohtaa missä palorypälehappo muuttuu aktivoiduksi etikkahaposki ja miten toisaalta käynnistyy sirtuunahapposykli ja beta-oksidaatio.
Eilen tuli hassu ajatus mieleen,. Nyt kirjoitan ajatuksen. Olen vuodesta toiseen katsonut sitä rinkuloitten hyvää ketjua fedox reaktiossa Se on kuin katsoisi oikein hyvän vaihdepyörän monipuolista vaihteistoa, , mutta siis vain vaihteistoa - näkemättä varsinaisesti muuta pyörää, kehitä tai pinnoja tai ylipäätänsä miten pyörää käytetään hyödyksi. Siis vaihteiston komeuden ympärillä oleva maisema on alunperin ollut kirjan valkoinen kartta, johon vuosi vuodelta on tullut jokin lisä kuin koriste joulukuuseen. Tiede on hurjasti edistynyt juuri elementaaristen tyhjien karttaläikkien kohdilta ja jos on tarkkana voisi vuosi vuodelta täydentää karttoja. Itseasiassa biologiset kartat ovat kuin ääretön palapeli.
Nyt sitten nämä sirtuiinit. Niitä on moneksi. Esim jotkut ovat mitokonriassa ja jotkut sytoplasmassa.
Jos ne kuitenkin toimivat deasetylaasina (etikkahapon irrottajana) kummassakin paikassa reaktioissa on jotan tyyppieroa tai tuotteen jatkokohtaloss on eroa, itseasiasas aitiollista hienosäätöä.
Katselin karttoja sirtuiinin osuudesta mitokondriaalisen etikkahapon aktivoimiseen. Asetylaattisynteesientsyymi (AceSC) muuttuu inaktiiviksi, jos siihen liittyy etikkahappo. Sirtuiini poistaa etikkahapon 8asetylaatin) ja synteesientsyymi aktivoituu jälleen ja samalla muut läsnäoleva tekijät jo aktivoivat etikkahapon (AcetylCoA muotoon).
Siis ynteeesientyymi löytää niitä tavallisia etikkahappoja , saa kaiekti joltain entsyymiltä avustuta otaa vastaan etikkahappo - Siten se on vaininaktiivi ja odottaa seuraavaa vaihetta. Sirtuiini tarkentaa etikkahapon jatkotalletustien ja ohjailee sitä täten kanavaan, joka johtaa aktivoituun muotoon ja sillehän on lukemattomasti käyttöä kehossa. Tämä kahden hiilen yksikön tallentaminen hyödyksi otaa energiaa (ATP). Etikkahappohan voi energeettömämin kondesoitua ketoaineeksi asetoetikkahapoksi ja vedeksi.
Tämä oli mieletäni hieno löytö sirtuiineista. Se sirtuiini, joka tekee tätä asiaa sytoplasmassa sallii tuottaa sellaista aktivoitua etikkahappoa, joka voi mennä rasva- ja steroidisynteesiin tai sirtuunahappokirtoon sytoplasmassa. Se sirtuiini, joka tekee aktivoitua asetaattia sisällä mitokondriassa vaikuttaa tehoa sitruunahapposykliin, jota tuottaa kolmin verroin enemmän hiilidioksidia (CO2) energian ja veden tuotannon sivutuotteen. Yleensähän rasva-aineperäiset AcetylCoA.t tuottavat irtoavan hiilidioksidin ja hiilihydraattiperäiset ovat sitruunahappokierron rakennemolekyylin komponetteja.
Tässä oli siis yksi hyvä uusi näkyväksi muuttunut sykli.
Mutta miksi otin tämän fytiiniblogiin?
Siksi, että Sirtuiinin deasetylaasifunktio on riippuvainen NAD+ koentsyymistä.
http://www.pnas.org/content/103/27/10230
NAD+ on nikotinamidi- adeniini-dinukleotidi . (Nikotinamidia muodostuu ravinnon tryptofaanihaposta kinolinaatti- ja nikotinaatti välimuotojen kautta).
Kun jokin proteiini pitää deasetyloida ( poistaa siitä etikkahappoa), sirtuiini katalysoi etikkahapon siirtymisen adenosylriboosiin ja muodostuu O-AAADPR ( O-asetyyli--ADP--riboosi) ja nikotinamidia (NAM) vapautuu.
Etikkahapon ( acetyl) vapauduttua jää ADP-riboosia, tästäkin jää lopulta ribosyyli-5-fosfaattia (R-5-P), joka on myös glykolyyttisen kartan tavallinen pentoosisokeri, jolle on monta käyttöä.
Kehossa on monta entsyymiä, jotka tekevät fosforiboosipyrofosfaattia PRPP- silläkin on monta käyttöä( puriinisynteesiin adeniiniin, guaniiniin, urasiiliin orottihappoon , siis AMP, GMP UMP tiehen).
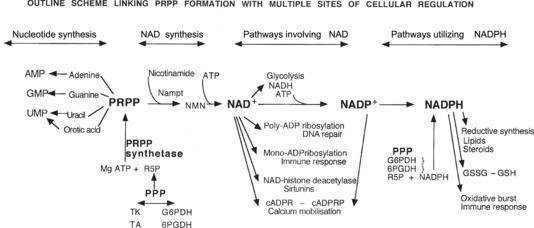
Mutta PRPP käyttyy myös NAD+ palauttamiseen.
https://www.google.com/search?q=phoshphoribosyl+pyrophosphate+synthesis&client=firefox-b&source=lnms&tbm=isch&sa=X&ved=0ahUKEwjTv77245bcAhUhCpoKHVeRChkQ_AUICigB&biw=1024&bih=471&dpr=1.88#imgrc=ieoB9En21_wXFM:
Nikotinamiditransferaasi siirtää NAM (nikotinamidin) PRPP:lle ja muodostuu NMN NikotinamidiNukleotidia.
ATP avustaa saamaan siitä takaisin sen koentsyymin NAD+, jota sirtuiini jälleen voi käyttää avuksi funktioonsa.
Tässä järjrstelmässä on tietty edullinen vaikutuksensa ihmisen aktiivilla energian käytöllä tämän deasetyloimissyklin aloittajana ( ja mitokondriahengityksen stimuloijana) eikä liika energia-aineen saanti (syönti) ilman tasapainoisesti vastaavaa energian käyttöä ole eduksi tälle hienosäädölle.
Vaikka fosfaatti on tässä syklissä paljolti sidottuna sokeriyhdisteeseen, muodostuu kuitenkin reaktioketjusta vapaata pyrofosfaattia PPi . Silloin muodostuva epäorgaaninen fosfaatti joutuu yleisfosfaattialtaan vaikutukseen.
Tässä toisaalta on toinen fosfaattiverkosto ( spesifisiä fosfaattiverkostoja on paljon), tässä tarkoitan inositolifosfaatti ja inositolipyrofosfaatteja. Häriöitä voi olla tietysti kummassakin sokerifosfaattiverkostossa.
Inositoliverkosto on lisäksi riippuvainen dietäärisesti inositolifosfaatin saannista, joka on orgaanista fosfaatia.
NAD+ taas on riippuvainen nikotiinihapon saannista ja sen muokkaamisesta koentsyymiksi, mikä on entsymaattinen tapahtuma. Nikotiinihappo onkin katsottu vitamiiniksi eikä ole luotettu siihen että kehokoneisto tryptofaanista tekee sitä riittävästi. matkan varrella on myös virhepolkuja kuten kinolinaatin kertyminen.
Ja kolmas asia, monissa vakavissa ihmiskuntataudeissa on jokin fosfaattiaineenvaihdunnan häiriö taustakartalla havaittavissa.
Tänään ajatellin löytää jotain yhteistä nimittäjää fosforibosyylipyrofosfaatille PRPP ja insositolipyrofosfaateille ja pyrofosfaatille PPi.
Sekä PRPP että InsP7 ovat kehon orgaanisia fosfaattimuotoja, sokeriin liittyneitä ja
PPi taas on epäorgaaninen muoto.
https://www.researchgate.net/profile/Glenda_Gillaspy/publication/273065355/figure/fig3/AS:271943068811285@1441847689019/Synthesis-of-inositol-pyrophosphate-Overview-of-the-Inositol-phosphate-pathway_Q320.jpg
{kind=link}
Siis: Jos pidän edessä PRPP karttaa ja toisaalta insositolifosfaatien kartaa. ovatko nämä aitioituneita
ja mistä kohtaa on interaktiot.
Otan esiin artikkeleita:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4291286/
- Why so many phosphates?
- https://www.researchgate.net/publication/229064431_How_inositol_pyrophosphates_control_cellular_phosphate_homeostasis
Abstract
Phosphorus in his phosphate PO(4)(3-) configuration is an essential constituent of all life forms. Phosphate diesters are at the core of nucleic acid structure, while phosphate monoester transmits information under the control of protein kinases and phosphatases. Due to these fundamental roles in biology it is not a surprise that phosphate cellular homeostasis is under tight control. Inositol pyrophosphates are organic molecules with the highest proportion of phosphate groups, and they are capable of regulating many biological processes, possibly by controlling energetic metabolism and adenosine triphosphate (ATP) production. Furthermore, inositol pyrophosphates influence inorganic polyphosphates (polyP) synthesis. The polymer polyP is solely constituted by phosphate groups and beside other known functions, it also plays a role in buffering cellular free phosphate [Pi] levels, an event that is ultimately necessary to generate ATP and inositol pyrophosphate. Although it is not yet clear how inositol pyrophosphates regulate cellular metabolism, understanding how inositol pyrophosphates influence phosphates homeostasis will help to clarify this important link. In this review I will describe the recent literature on this topic, with in the hope of inspiring further research in this fascinating area of biology.
(PDF) How inositol pyrophosphates control cellular phosphate homeostasis?. Available from: https://www.researchgate.net/publication/229064431_How_inositol_pyrophosphates_control_cellular_phosphate_homeostasis [accessed Jul 11 2018].
Entä PRPP liika syntetisoituminen?
https://www.ncbi.nlm.nih.gov/pubmed/20301734
Inga kommentarer:
Skicka en kommentar