
(Vaikuttaa siltä että IP6K2 /IP7 tehostaa kaspaasi-8 kautta tapahtuvaa apoptoosia.
Tässä kuvaan tavallisen kertomuksen kaspaaseista lähteenä Dionisios Chrysiksen väitöskirjan taustateksti. The Role of apoptosis in growth plate cartilage during normal and abnormal growth. STH 2004. ISBN 91-628-6293-6.)
Kaspaasit osavaikuttajina apoptoosissa
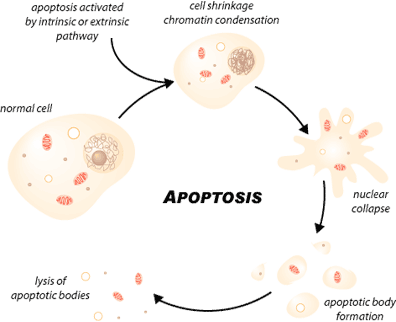
APOPTOOSI on fysiologinen tapahtuma, jossa solu katoaa tulehduksettomasti
Sellainen solukuolema on ohjelmoitua, missä on erityiset morfologiset ja biokemialliset piirteet, kuten tumakromatiinin tiivistyminen ja DNA:n fragmentoituminen, solun tilavuuden vähenemä ja plasmakalvon muuntumiset, apoptosomien muodostuminen. Näistä seikoista tunnistetaan apoptoosi solun ulkopuolelta käsin ja sellainen solu fagosytoituu. Tämä on fysiologinen prosessi, joka on tärkeä solujen normaalikasvulle ja homeostaasille monisoluisissa elimissä ( Kaikki yhden puolesta ja yksi kaikkien puolesta –periaatteella). Myös apoptoosi on tässä tärkeä geenisäätöinen tekijä yksittäisen orgaanin ja täten koko henkilön kannalta.
Immuunisysteemin säätelyssä tarvitaan apoptoosikykyä.
Tumorogeneesissä on apoptoosilla osuus.
SOLUN GEENISTÖ kantaa apoptoosikykyä.
Miten solu voi apoptoitua?
Se järjestää ohjelman, joka toimii loppuun asti senkin jälkeen, kun solun geenistö on hajautunut. Geneettisesti koodattu ohjelma on evoluutiossa hyvin säilyneenä soluissa ja johtaa tarvittaessa yksityisen solun tehokkaaseen, loogiseen ja siistiin apoptoitumiseen. Solu ikäänkuin ”haihtuu olemattomiin”- Esim vanhetessa keho ikään kuin näkymättömästi kutistuu kaikilta kudoksiltaan fysiologisesti ja loogisesti kuitenkin yksilön elinkyvyn ja tahdonalaisen suuntauksen hyväksi. Tahdonalainen aivotoiminta on korkein määrääjä, arvelen.
APOPTOOTTINEN KASKADI
Apotoosi on jaettavissa kolmeen päävaiheeseen
• Aloitusvaihe, initiaatio
• Exekutiivinen vaihe, lopullinen päätös apoptoitumisesta tehdään.
• Resolutiivinen vaihe, lopputapahtumat.
1. APOPTOOSIN ALKAA jokin seikka, triggeri, kuten kasvutekijän puuttuminen tai vähäisyys, deprivaatio, kuolonreseptoriin sitoutunut ligandi tai genotoksisen tekijän vaikutus. Nämä aiheuttavat, että solu havaitsee elimistölle ja genomille parhaimmaksi ratkaisuksi katoamisen soluyhteiskunnasta. Yksityisen solun genomi asentuu apoptoosiin. Apoptoosin merkitys on integroituna loppusummana kuitenkin globaalin genomin paras mahdollinen elossapysyminen. Apoptoosissa on logiikkaa. Esim. ihminen vanhetessaan pienenee kaikin tavoin ja saattaa täten säilyttää pirteän henkisen struktuurin ja elämänlaadun ja liikkuvuuden ja tarve liikkumiseen /liikunta pitää yllä lokomotorista struktuuria feed backin omaisesti.
2. PÄÄTÖS APOPTOITUMISESTA, Kun exekutiivinen vaihe on tapahtunut, siinä solun genomi alkaa hajota, eikä solu sen jälkeen voi päättää mitään toista muutosta. Spesifiset proteaasit ja nukleaasit hajoittavat rakenteen ja signaloivat proteiinit sekä geneettisen materiaalin johtaen erityisiin morfologisiin muutoksiin.
3. RESOLUUTIOVAIHEESSA solupinta antaa hätämerkkiä lähistöllä oleville terveille soluille, joten ne havaitsevat tämän solun fagosytoosivalmiuden merkit ja niin ne fagosytoivat sen jäänteet. Tämä varmistaa, että mitään itsestään kasvavaa geneettistä materiaalia ei jää jäljelle, sillä geneettinen materiaali on itseään kopioivaa, varsinkin kun se on elävän kehon sisällä, lähellä toisia soluja, jonne se voisi translokoitua. Siksi apoptoosin täydellinen onnistuminen on fysiologisesti ottaen soluyhteiskunnan terveyttä.
MISTÄ TUNNISTAA APOPTOOSIVAIHEITA LÄPIKÄYVÄN SOLUN
Näillä soluilla on morfologisia ja biokemiallisia piirteitä. Nähdään muutos solukalvon integriteetissä, kromatiinia aggregoituu, tuma ja sytoplasma kondensoituu ja tapahtuu sytoplasman ja tuman erkanemista kalvojen ympäröimiksi rakkuloiksi, apoptosomeiksi, joissa on sisällä ribosomeja, morfologisesti ehjiä mitokondrioita ja tumamateriaalia. Solu integroi ja tarkentaa apoptoosinsa asettaen nämä geneettisesti aktiivit arat ainekset erikseen paketoituna odottamaan jatkokäsittelyä. Koeputkessa nämä apoptoosipaketit tunnistetaan nopeasti ja makrofagit alkavat fagosytoida niitä tai lähellä oleva epiteelikin voi toimia fagosytoijana.
Tähän apoptoottisten solujen tehokkaaseen poistamiseen in vivo ei tarvitse aktivoida tulehdusvastetta. Asia hoituu ikäänkuin sopusoinnissa lähisolujen kanssa, missä kaikille on hyödyksi tulehdukseton miljöö.
http://www.scq.ubc.ca/wp-content/uploads/2006/07/Apoptosis.gif
APOPTOOSITIEN KOMPONENTIT
Miten tällainen hieno järjestelmä toimii?
Nisäkäskehossa tapahtuvan apoptoosin kaikkia yksityiskohtia ei ole selvitetty , mutta on voitu todeta yhtäläisyyksiä nematoda-solujen apoptoosiin. Kuolonsäätäjiä on näissä ainakin kaksi pääluokkaa:
• Onkogeenien perhe
• Cysteiiniproteaasien perhe, kaspaasit
ONKOGEENIEN PERHE, Bcl-2
Prototyyppinä on Bcl-2. Ryhmään kuuluu Bcl-2 ja Bax.
ANTIAPOPTOOTTISIA eli kuolon antagonisteja, vastavaikuttajia, ovat Bcl-2, Bcl-x1, Bcl-w, Mcl-1, Bfl-1, Brag.
PROAPOPTOOTTISIA eli kuolon agonisteja, myötävaikuttajia, ovat Bax, Bak, Bcl-xS, Bik.
http://www.ncbi.nlm.nih.gov/bookshelf/picrender.fcgi?book=eurekah∂=A8980&blobname=ch736f2.gif
CYSTEIINIPROTEAASIEN PERHE , Caspases
ICE-Like molecules, caspases
Interleukiini 1-beta (IL-1b) converting enzyme like molecules
Näiden toinen nimi siis on kaspaasit. Kaspaasit ovat apoptoosin effektoreja ja vastuussa proteiinien spesifisten tunnistuskohtien proteolyyttisesta pilkkoamisesta. Pääasiassa ne samalla inaktivoivat kohdeproteiinia. Tosin on muutamia tapauksia, että ne aktivoivat proteiinifunktion.
Mitä proteiineja kaspaasit pilkkovat?
Kaspaasit hajoittavat niitä proteiineja, jotka ovat osallistuneet olennaisina komponentteina sytoplasman ja tuman struktuurien ylläpitämiseen. Tällaisia ovat lamiini A ja fodriini. DNA-korjausjärjestelmän jäsenet ovat niitä myös, kuten PARP ( poly-ADP-riboosi-polymeraasi)
DNA:n hajoamisen estäjät ovat myös niitä: Tällainen on CAD-estäjä, kaspaasiaktivoidun desokxyribonukleaasin estäjä.
Solusyklin säätelyyn osallistuvat proteiinit ovat myös kaspaasien kohteina, kuten esim pRb( retinoblastoomaproteiini)
• Initiaattorikaspaasit (-8, -9)
• Exekutoriset kaspaasit (-3,-6,-7)
Normaalisti kaspaasit ovat inaktiiveina proentsyymeinä läsnä solussa ja ne tarvitsevat sen takia aktivoituakseen proteolyyttisen prosessoitumisen. Aktivoituminen tapahtuu joko autokatalyyttisesti tai toisten kaspaasien vaikutuksesta.
Apoptoosin aloittajakaspaasit, initiaattorit ovat kaspaasi-8 ja kaspaasi-9
Näillä aktivoituja muita kaspaaseja kutsutaan exekutorisiksi kaspaaseiksi kuten kaspaasi-3, kaspaasi-6 ja kaspaasi-7.
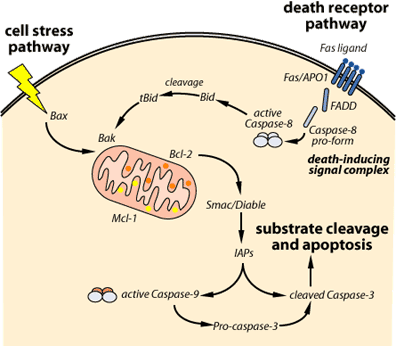
Solun ulkoiset ja sisäiset APOPTOOTTISET TIET
Vaikka apoptoosin alkamisen tarkkoja mekanismeja ei tunnetakaan in vivo, tiedetään, että vasteen kehittyminen on riippuvainen triggeröivästä tekijästä ja on solutyypille spesifinen.
Apoptoosin signaalitransduktio saattanee edetä kahdella toisistaan eriävällä reitillä.
• reseptorivälitteinen eli ulkoinen apoptoositie EXTRINSIC
• mitokondriavälitteinen eli sisäinen apoptoositie, INTRINSIC.
ULKOTIE (EXTRINSIC ) reseptorivälitteinen apoptoositie
Tämä tie vaatii ligandin(L) sitoutumisen kuolonreseptoriin solun pinnalla.
Esimerkiksi TNF( tuumorinekroositekijä) sitoutuu reseptoriin TNF-1R ja rekrytoi kaksi signaalitransduktiomolekyyliä
1. TNF-1R –assosioituneen kuolondomaanin(death domain, DD) proteiinin , TRADD
2. Fas-assosioituneen proteiinin, jolla myös on kuolondomaani, FADD.
TNFR-perheen kuolonreseptoreihin DR kuuluu TNFR1, Fas(CD95), DR3/WSL sekä
TNF-kaltainen apoptoosia indusoiva ligandi TRAIL/Apo21 reseptori (TRAIL-R1/DR4, TRAIL-R2/DR5).
Tämän ryhmän jäsenille on luontaista, että niillä on 2-5 cysteiinirikasta extrasellulaarista toistojaksoa. Kuolonreseptoreilla on myös solunsisäinen DD-domaanin nimellä kutsuttu aminohappojakso C-terminaalissa.
Kun nämä kuolonreseptorit ovat sitoutuneet vastaaviin ligandeihin (TNF tai lymfotoksiini, FasL, DR3-ligandi tai TRAIL/Apo-21), apoptoosi voi käynnistyä.
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah∂=A8980&rendertype=figure&id=A8987
Kaksi prokaspaasi-8-molekyyliä voi prosessoida toisensa aktiiviin muotoon. Aktiivi kaspaasi-8 voi siten triggeröidä päälle apoptoottisen kaspaasikaskadin.
(Uutta tietoa on , että PP—IP5 ja IP6K2 vaikuttavat kaspaasi-8 teitse apoptoosia edistävästi).
SISÄTIE eli mitokondriaalinen apoptoositie
Tieto lisääntyy mitokondrian essentiellistä osuudesta useissa apoptoosimuodoissa, koska se vapauttaa apoptogeenisiä tekijöitä kuten sytokromi C- molekyylin (Cyt C) intermembraanitilasta sytoplasmaan ja täten aktivoi apoptoosin ”downstream execution” tapahtumat. Cyt c irtoama solun energialaitoksesta, mitokondriasta, on kuin mitokondrian hengen poistuminen.
http://www.ncbi.nlm.nih.gov/bookshelf/picrender.fcgi?book=eurekah∂=A32938&blobname=ch174f3.jp
http://www.reading.ac.uk/nitricoxide/intro/apoptosis/mito.htm
Antiapoptoottiset Bcl-2-proteiiniperheen jäsenet koettavat pääasiassa estää mitokondriaalisia muutoksia ja täten mitokondriasta johtuvaa apoptoosia. Nämä antiapoptoottiset Bcl-2 ja Bcl-X1 sijaitsevat mitokondrian ulkokalvossa.
Sitävastoin proapoptoottiset jäsenet sijaitsevat joko sytosolissa tai mitokondrian kalvossa. Ryhmät vastavaikuttavat toisiinsa. Antiapoptoottiset koettavat ehkäistä sytokromi C:n irtoamista ja täten pitkittää solun pysymistä hengissä. Mutta proapoptoottiset aloittavat cyt c-hengitysketjutekijän irrottamisella mitokondriasta. Tämä taas aktivoi executorisia kaspaaseja muodostamalla apoptosomin.
Apoptosomin muodostaa cyt c, proteaasia aktivoiva tekijä-1 Apaf-1 ja prokaspaasi-9.
http://www.reading.ac.uk/nitricoxide/intro/apoptosis/mito2.jpg
On toinenkin mitokondriaalinen kaspaasiaktivaattori.
Tämän nimi on Smac/DIABLO, ja sitä vapautuu mitokondriasta c-sytokromin mukana. Se edistää kaspaasiaktivaatiota liittymällä apoptosomiin ja estämällä apoptoosin inhibiittoriproteiinia IAP.
http://www.rsc.org/ej/NP/2001/a909080k/a909080k-f6.gif
Smac/DIABLO sitoutuu IAP-proteiiniin ja rikkoo sen liittymisen prosessoituvaan kaspaasi-9-molekyyliin ja niin se sallii kaspaasi-9- entsyymin prosessoitua edelleen ja aktivoida kaspaasi-3-proteiinin, joka on executorisia kaspaaseja.
http://www.scq.ubc.ca/wp-content/uploads/2006/07/apoptosispathways.gif
KASPAASEISTA RIIPPUMATTOMAT APOPTOOSITIET:
On osoitusta siitä, että tapahtuu apoptooseja ilman kaspaasien osuutta. Nimittäin vaikka kaspaasit inhiboitaisiin, tapahtuu silti solun kuolemaa ja sen merkkinä tuman kondensaatiota, kalvojen kuplimisia ja solujen kutistumisia. Tästä on päätelty, että kaspaaseja ei tarvita apoptoosin kaikkiin aspekteihin.
AIF
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah∂=A32938&rendertype=figure&id=A32956
Yksi avainmolekyyli kaspaaseista riippumattomaan solukuolemaan on apoptoosia indusoiva tekijä AIF (Apoptosis Inducing Factor). Solukuolon signaloinnissa AIF translokoituu mitokondriaalisesta intermembraanitilasta tumaan, missä se sitoutuu DNA:han ja aiheuttaa kaspaaseista riippumattoman kromatiinin kondensaation .
5.6.2009 17:39
KOMMENTTINI: IP7 muodon tulee pystyä muuttua IP8 muodoksi, muuten se jää apotoosia stimuloivaksi kun taas IP8 muoto antaa suojaa kromosomille.
fredag 5 juni 2009
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Syöpälääkkeet koettavat käyttää kalsiumpitoisuutta työvälineenä
Syöpälääkkeet vaikuttavat kalsiumin signalointijärjestelmän kautta tai ainakin koettavat vaikuttaa.
Fosfoinositidien ja inositolifosfaattien laaja aineenvaihduntakartta säätelee kalsiumin signalointia terminaalisena aseena niin energian hankkimisessa kuin solujen muokkaamisessa, erityksissä ja loppuvaoheessa vanhojen solujen hävityksissä uusien solujen tieltä.
LÄHDE: Hägg Olofsson Maria. Translational studies of drug-induced tumor cell death. Karolinska insitutet. 2006. ISBN 91-7140-855-X.
Suomennosta ja välillä kommentteja.
On tärkeää ymmärtää syövänvastaisten lääkkeitten mekanismia, jotta voisi saada tuloksia syövän hoidosta. Tutkija on tässä tutkimuksessaan käyttänyt kliinisessä käytössä olevaa anti-kanserogeeniterapian ainetta ja muitakin molekyylejä tutkiessaan tuumorisolun tuhoutumista. Mainitaan cisplatiini ja elliptisiini. Väitöskirjatyössä mm. verrattiin estramustiinia, vinorelbiinia ja docetakselia.
CISPLATIINI on tärkeä agenssi eri maligniteettimuotojen hoidossa ja sillä havaitaan olevan monimukaisempaa toiminnallista vaikutusta, mitä aiemmin on arveltukaan. Eri annosten cisplatiinia havaitaan indusoivan kahta erilaista soluvastetta.
http://tbn2.google.com/images?q=tbn:94j-3BW5HcYAKM:
http://dic.academic.ru/pictures/enwiki/67/Cisplatin-2D.png
http://www.rbej.com/content/figures/1477-7827-1-66-2.jpg
Matalista annoksista tulee esiin ”senescence”, solun vanhentumismuutoksia.
Suurista annoksista cisplatiinia alkaa ”apoptosis” , ohjelmoitu solukuolema.
Mekanismeina näissä on (mm) vapaitten happiradikaalien muodostuminen ( reactive oxygen species, ROS) ja kalsiumin (Ca++) vaikutus. Näitä vasteita triggeröi esiin erilaiset signaalitekijät. Apoptoosi triggeröityy esiin sytoplasmisilla mekanismeilla ja ”senescence”- solun vanhentumisilmiö, DNA-vaurioista.
ELLIPTISIINI indusoi stressiproteiinin HSP70.
http://www.chemicalregister.com/upload/cr/519-23-3.png
(5,11-dimethylpyrido[4,3-b]carbazole)
Kasvisperäisen alkaloidin ELLIPTISIINI, havaittiin olevan cisplatiini- molekyylille synergistinen aine (Cisplatiini aiheuttaa myös endoplasmisen retikulumin (ER) stressin). Yksityiskohtaisissa tutkimuksissa todettiin, että elliptisiini indusoi soluorganellitasossa endoplasmisen verkoston varhaisia stressivasteita, joita luonnehtii endoplasmisen chaperone-proteiinin GRP78/BiP muodostuminen stressivasteena ( ennen kuin sytokromi c pilkkoutui esiin ja CK18 ilmeni) ja XBP1 transkriptiotekijää esiinpilkkoutuma .
Netistä löytyy maininta:
(GRP78/BiP Is Required for Cell Proliferation and Protecting the ...
ER stress induction of GRP78/BiP represents a major prosurvival arm of the unfolded protein response (UPR)).
( The 78 kDa glucose regulated protein/BiP (GRP78) belongs to the family of ~70 kDa heat shock proteins (HSP 70))
Stressiproteiinit HSP koettavat laskostaa uudelleen denaturoituja proteiineja ja täten pitää solua hengissä.
GRP on glukoosia säätelevä protein, Glucose Regulating Protein. GRP78 on endoplasmisen retikulumin eräs ”chaperone ", HSP70.
http://upload.wikimedia.org/wikipedia/en/2/21/PROTEIN_FOLDING_SIMPLIFIED_JPEG_small.jpg
Tässä väitöskirjassa esitetyt cisplatiinin ja elliptisiinin tutkimukset esittävät monen anticancerogeenin lääkkeen kompleksista vaikutusmallia.
MITÄ pro ja anti -APOPTOOSITEKIJITÄ ON SOLUORGANELLITASOSSA?
Endoplasmisessa retikulumissa sijaitsee Bcl-2-perheen proteiineja ja Bcl-2 itse on endoplasmiselle verkostolle sytoprotektiivinen, lojaali tekijä perheenjäsenineen.
http://journals.prous.com/journals/dot/20003607/html/dt360415/images/Alz_f29.gif
Lysosomien katepsiinit taas indusoivat apoptoosia TNF tekijän kautta.
Jotkut syöpälääkkeet taas kohdistavat vaikutuksen mitokondriaan suoraan.
Nämä organellit, ER ja lysosomit, voivat herkistyä stressille ja välittää signaalit mitokondria-organellille, johon tulee reikiä ja mitokondria joutuu apoptoosiin
ER herkistyy stressille stressiproteiiniensa (HSP) , chaperoniensa kautta, kalsiumia sitovien proteiiniensa ja kalsiumia vapauttavien kanaviensa välityksllä ja siitä välittyy ER-vasteet mitokondriaan.
SOLUTASO:
Tuman DNA vaurio ja DR-reseptorien ( death receptors) triggeröityminen ovat tavalliset ulkoiset apoptoosisignaalit solulle), esim gamma ja röntgensäteilyvaikutuksesta.
http://www.promega.com/paguide/images/1628MC10_7A.jpg
KALSIUMIN (Ca++) signaloinnin on osoitettu olevan taustatyövälineenä monen vaikuttavan aineen aihettamassa apoptoosissa. Niin myös cisplatiinilla. Jotta voitiin paremmin osoittaa kalsiumin signaloinnin ratkaisevaa osuutta, tutkittiin 40 eri yhdisteen aiheuttamaa apoptoosia ja nämä aineet ovat seulottavissa esiin kemiallisesta kirjastosta. Näistä tutkimuksista voidaan päätellä, että KALPAIINI ( calpain) ja KALMODULIINIKINAASI II (CaMKII) ovat tärkeitä tekijöitä kalsiumin signaloinnin välittämisessä apoptoositapahtumassa.
• Kalsiumin osuus apoptoottisessa prosessissa oli myöhäinen ja kalsiumin signalointi havaittiin taustatekijänä Jun N-terminaalisen kinaasin (JNK) aktivaatiossa.
http://journals.prous.com/journals/dnp/20041710/html/dn170646/images/Li_f1.gif
(KOMMENTTI: PI3K sanotaan survival tekijäksi. Sen sijaan IP6K2 on apoptoosin tekijä, mikä on uudempaa tietoa) http://ccforum.com/content/figures/cc2387-1-l.jpg
Toinen väitöskirjatyön päämäärä oli kehittää monitorointimetodia in vivo-tarkoituksiin syöpäterapialääkkeitten aktiivisuuden mittauksia varten.
KASPAASIEN OSUUS apoptoosiaktiviteetin toimittajana. (Tästä kirjoitan erikseen)
Tutkijat mittasivat kaspaasien (CASPASE) pilkkomien SYTOKERATIINI 18-fragmenttien (CK18) määrää potilaan seerumista syöpäterapian aikana. Metodista kehkeytyi kiinnostavaa kliinistä informaatiota.
Syöpälääke DOCETAXEL(x) (http://en.wikipedia.org/wiki/Docetaxel) havaittiin tehokkaammaksi tuumorin apoptoosin indusoijaksi prostatasyövässä verrattuna kahteen muuhun tutkittuun valmisteesen. Eräs erityinen mielenkiintoinen havainto oli se, että apoptoosi indusoitui esiin useitten hoitojaksojen kuluessa.
Kun mitattiin kaspaasilla esiin pilkkoutuneitten totaalisytokeratiini-18 fraktion määriä seerumista, saatiin osoitusta siitä, että
CEF- kombinaatio, jota käytettiin rintasyövän hoitoon, indusoi pääsiassa nekroottisen solutuhovasteen.(CEF= syklofosfamidi, epirubisiini, 5-fluorourasiili). http://www.germanbreastgroup.de/nnbc3/english.html
Kun parantuu metodit, joilla määritetään lääkevaikutuksia in vivo olosuhteissa on odotettavissa uusien lääkkeiden kliiniseen kehityksen apua ja samoin hyötyä kliinisen työhön.
(x) Docetaxel on semisynteettinen taksaani, kemoterapeuttinen aine, joka sitoo beeta-tubuliinia, stabiloi mikrotubuluksia ja aiheuttaa solusyklin pysähtymää (CELL CYCLE ARREST) ja apoptoosia (2001). Jos tällä käsiteltiin 3 viikon välein, tulokset hengissäpysymisen ja paranemiseen suhteen paremmat kuin mitoxantronilla (2004).
(Kommentti: Väitöskirjassa kuvattuja lääkkeitä ovat myös
Estramustiini polyfosfaatti (EMP) on (nornitrogen mustard)-estradiolikonjugaatti, joka vaikuttaa mikrotubulaariseen dynamiikkaan ( 1999). Sitä on käytetty prostatasyöpään vuosia.
http://en.wikipedia.org/wiki/Estramustine
Vinorelbine vaikuttaa myös mikrotubuluksiin ja sillä on aktiivisuutta prostatasyöpää vastaan.
http://en.wikipedia.org/wiki/Vinorelbine
Väitöskirjatyössä verrattiin estramustiinia, vinorelbiinia ja docetakselia).
VAIKUTUSMEKANISMIN TAUSTASTA KALSIUMIN SUHTEEN
( On paljon strategioita, joita koetetaan kayttää, mutta monsisa konvergoituu viime käden tärkeäksi signaloivaksi vaikuttajaksi Calsium joni ja täten inositolifosfaatti järjestelmän molekylääriset resurssit).
http://www.nature.com/cdd/journal/v13/n8/fig_tab/4401940f3.html#figure-title)
Väitöskirjatyöhön liittyy kuva KALSIUMIN SIGNALOINNISTA TÄSSÄ YHTEYDESSÄ. Siihen kuuluvia osatekijöitä ovat mm:
CALPAIN , Ca++aktivoituva proteaasi kalpaiini on tärkeä apoptoosin signaalivälittäjä.
http://nashua.case.edu/pathwaysweb/GOPathwaysAnalysisTool/resource/GO/images/0004198.png
Kalpaiini vaikuttaa eräänlaiselta solun serubbaabelilta, (ett slags härjande på fältet).
www.nature.com/.../v28/n4/fig_tab/9600595f2.html
CALPEPTIN, kalpeptiini on kalpaiinin inhibiittori ja voi estää 19 yhdistettä niistä 40 tutkitusta apoptoosia aiheuttavasta aineesta.
Kalpaiini ei yksinään kuitenkaan voi vastata kaikista apoptoosivaikutuksista.
CALMODULIN (CaM), kalmoduliini on tärkeä ja pääasiallinen intrasellulaarinen Ca++-sitova proteiini ja se voi olla agonisti tai antagonisti apoptoosille.
Jos W7- tekijällä estettiin kalmoduliini, apoptoosia aiheuttavista 40 tekijästä estyi 17 tekijää. Kalmoduliinin osuus kuitenkin oli tärkeä apoptoosissa.
http://www.nature.com/nrn/journal/v6/n4/images/nrn1647-f2.jpg
CALCINEURIN (proteiinifosfataasi 2B, PP2B), kalneuriini. Tämä on Ca-kalmoduliinilla aktivoituva seriini-treoniini PP ja aktivoituu solunsisäisestä kalsiumista (Ca++)i. Kalkineuriini poistaa fosfaattia ASK-1:stä, joka on apoptoosia signaloiva kinaasi-1 ja tällöin tämän kinaasin aktiviteetti pääsee lisääntymään.
ASK1 sitten aktivoi JNK ja p38 ja johtaa kohti apoptoosia.
Kalkineuriinia pidetään solukuoleman moduloijana. Se on myös sensitiivinen solustressille, vaikka tässä tapauksesa oksidaatio johtaa entsyymin inaktivaatioon. Mitään vahvaa apoptoosin estymistä ei siitä kuitenkaan saa aikaan inhibiittorilla.
(Kommentti: Stressiproteiineista se voi aktivoitua ja johtaa tällöin solun selviämiseen.
(http://molinterv.aspetjournals.org/cgi/content/full/2/5/284/F1
“The unfolded protein response (UPR) is a transcriptional and translational – tämä tekijä on otettava huomoon, sillä nykyajan ihmisellä on aina sen verran solustressejä- mikä sitten reaktioketjun loppusuoralla vaikuttaa solun apoptoosin ja eloonjäämisen exekuutiossa- asian voi miltei pelkistää IP3/ IP7 suhteeksi - ehkä myös sanoilla, kalsiumin säädelty vaimennus / kalsiumin progrediointiin sysätty runsas lisäys ).
USEITA KALMODULIINISTA riippuvia proteiineja on identifioitu ja yksi näistä on
CALMODULIN KINASE II ( CaMK II), joka toimii välittäjänä solun kalsiumin ja kalmoduliinivälitteisen apoptoosin kesken. CaMK II voi fosforyloida ASK-1 kinaasia, mutta suhde apoptoosiin ei ole tutkittu.
Kuitenkin CaMK II inhibiittori (KN-93) kykeni blokeeraamaan apoptoosin 21 tapauksessa 40 :stä.
JNK-välitteiselle apoptoosille löytyy useita signaalireittejä, kuten p53 ( ns. genomin suojelijamolekyyli) sekä Bak ja Bax.
http://genesdev.cshlp.org/content/13/8/916/F1.large.jpg
http://www-rcf.usc.edu/~forsburg/images/Image10.jpg
http://herkules.oulu.fi/isbn9514270398/html/graphic22.png
http://www.nature.com/bjc/journal/v94/n4/images/6602955f7.gif
JNK-välitteisen apopoosin pystyi estämään eräät aineet:kuten JNK-estäjä SP600125.
Kts HSP stressiproteiinien osuutta:
http://www.biolsci.org/v03/p0020/ijbsv03p0020g01.jpg
CALSIUMIN SIGNALOINTI aktivoi JNK:n - joissain tutkimuksissa. Osa signaaleista kulkee CaMK II / JNK tietä.Tutkija mainitsi, että ollaan tekemässä lisätutkimuksia, joissa selvitellään, onko JNK aktivaation edellä CaMKII vaikutus.
5.6.2009 11:18
Fosfoinositidien ja inositolifosfaattien laaja aineenvaihduntakartta säätelee kalsiumin signalointia terminaalisena aseena niin energian hankkimisessa kuin solujen muokkaamisessa, erityksissä ja loppuvaoheessa vanhojen solujen hävityksissä uusien solujen tieltä.
LÄHDE: Hägg Olofsson Maria. Translational studies of drug-induced tumor cell death. Karolinska insitutet. 2006. ISBN 91-7140-855-X.
Suomennosta ja välillä kommentteja.
On tärkeää ymmärtää syövänvastaisten lääkkeitten mekanismia, jotta voisi saada tuloksia syövän hoidosta. Tutkija on tässä tutkimuksessaan käyttänyt kliinisessä käytössä olevaa anti-kanserogeeniterapian ainetta ja muitakin molekyylejä tutkiessaan tuumorisolun tuhoutumista. Mainitaan cisplatiini ja elliptisiini. Väitöskirjatyössä mm. verrattiin estramustiinia, vinorelbiinia ja docetakselia.
CISPLATIINI on tärkeä agenssi eri maligniteettimuotojen hoidossa ja sillä havaitaan olevan monimukaisempaa toiminnallista vaikutusta, mitä aiemmin on arveltukaan. Eri annosten cisplatiinia havaitaan indusoivan kahta erilaista soluvastetta.
http://tbn2.google.com/images?q=tbn:94j-3BW5HcYAKM:
http://dic.academic.ru/pictures/enwiki/67/Cisplatin-2D.png
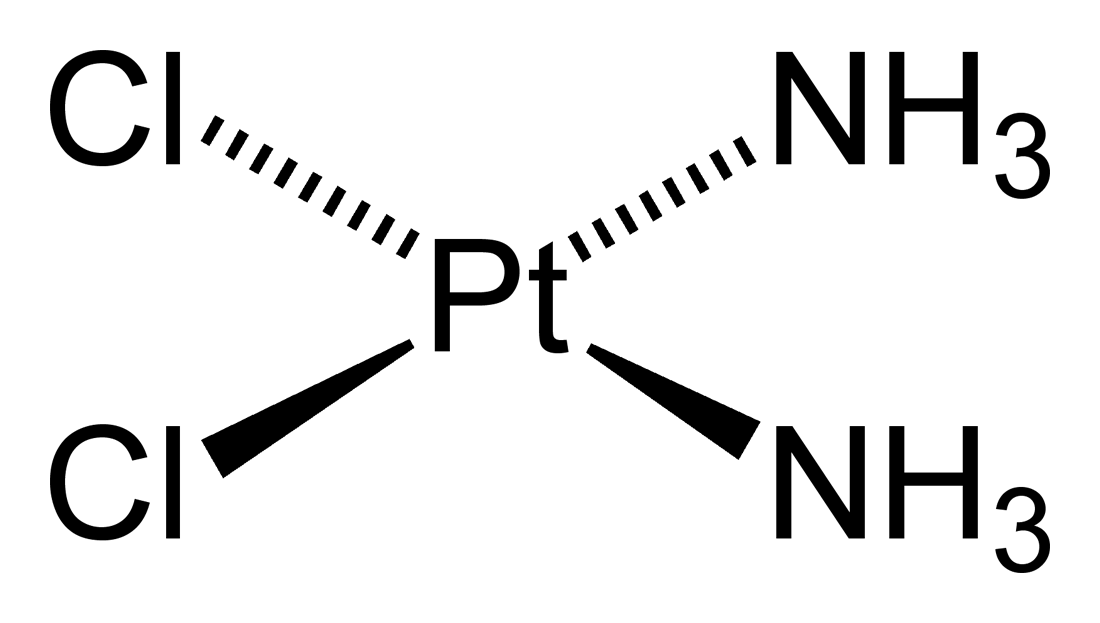{kind=link}
http://www.rbej.com/content/figures/1477-7827-1-66-2.jpg
{kind=link}
Matalista annoksista tulee esiin ”senescence”, solun vanhentumismuutoksia.
Suurista annoksista cisplatiinia alkaa ”apoptosis” , ohjelmoitu solukuolema.
Mekanismeina näissä on (mm) vapaitten happiradikaalien muodostuminen ( reactive oxygen species, ROS) ja kalsiumin (Ca++) vaikutus. Näitä vasteita triggeröi esiin erilaiset signaalitekijät. Apoptoosi triggeröityy esiin sytoplasmisilla mekanismeilla ja ”senescence”- solun vanhentumisilmiö, DNA-vaurioista.
ELLIPTISIINI indusoi stressiproteiinin HSP70.
http://www.chemicalregister.com/upload/cr/519-23-3.png
{kind=link}
(5,11-dimethylpyrido[4,3-b]carbazole)
Kasvisperäisen alkaloidin ELLIPTISIINI, havaittiin olevan cisplatiini- molekyylille synergistinen aine (Cisplatiini aiheuttaa myös endoplasmisen retikulumin (ER) stressin). Yksityiskohtaisissa tutkimuksissa todettiin, että elliptisiini indusoi soluorganellitasossa endoplasmisen verkoston varhaisia stressivasteita, joita luonnehtii endoplasmisen chaperone-proteiinin GRP78/BiP muodostuminen stressivasteena ( ennen kuin sytokromi c pilkkoutui esiin ja CK18 ilmeni) ja XBP1 transkriptiotekijää esiinpilkkoutuma .
Netistä löytyy maininta:
(GRP78/BiP Is Required for Cell Proliferation and Protecting the ...
ER stress induction of GRP78/BiP represents a major prosurvival arm of the unfolded protein response (UPR)).
( The 78 kDa glucose regulated protein/BiP (GRP78) belongs to the family of ~70 kDa heat shock proteins (HSP 70))
Stressiproteiinit HSP koettavat laskostaa uudelleen denaturoituja proteiineja ja täten pitää solua hengissä.
GRP on glukoosia säätelevä protein, Glucose Regulating Protein. GRP78 on endoplasmisen retikulumin eräs ”chaperone ", HSP70.
http://upload.wikimedia.org/wikipedia/en/2/21/PROTEIN_FOLDING_SIMPLIFIED_JPEG_small.jpg
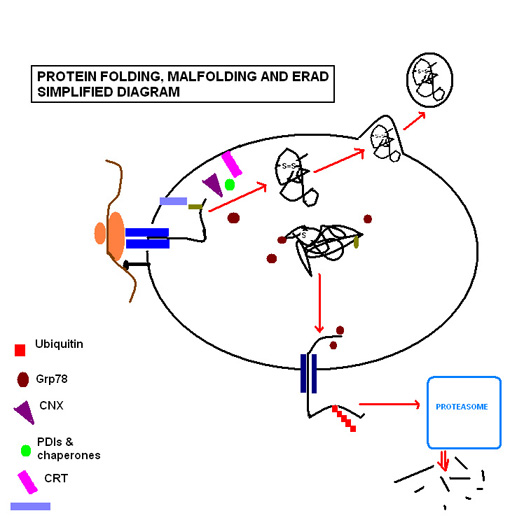{kind=link}
Tässä väitöskirjassa esitetyt cisplatiinin ja elliptisiinin tutkimukset esittävät monen anticancerogeenin lääkkeen kompleksista vaikutusmallia.
MITÄ pro ja anti -APOPTOOSITEKIJITÄ ON SOLUORGANELLITASOSSA?
Endoplasmisessa retikulumissa sijaitsee Bcl-2-perheen proteiineja ja Bcl-2 itse on endoplasmiselle verkostolle sytoprotektiivinen, lojaali tekijä perheenjäsenineen.
http://journals.prous.com/journals/dot/20003607/html/dt360415/images/Alz_f29.gif
{kind=link}
Lysosomien katepsiinit taas indusoivat apoptoosia TNF tekijän kautta.
Jotkut syöpälääkkeet taas kohdistavat vaikutuksen mitokondriaan suoraan.
Nämä organellit, ER ja lysosomit, voivat herkistyä stressille ja välittää signaalit mitokondria-organellille, johon tulee reikiä ja mitokondria joutuu apoptoosiin
ER herkistyy stressille stressiproteiiniensa (HSP) , chaperoniensa kautta, kalsiumia sitovien proteiiniensa ja kalsiumia vapauttavien kanaviensa välityksllä ja siitä välittyy ER-vasteet mitokondriaan.
SOLUTASO:
Tuman DNA vaurio ja DR-reseptorien ( death receptors) triggeröityminen ovat tavalliset ulkoiset apoptoosisignaalit solulle), esim gamma ja röntgensäteilyvaikutuksesta.
http://www.promega.com/paguide/images/1628MC10_7A.jpg
{kind=link}
KALSIUMIN (Ca++) signaloinnin on osoitettu olevan taustatyövälineenä monen vaikuttavan aineen aihettamassa apoptoosissa. Niin myös cisplatiinilla. Jotta voitiin paremmin osoittaa kalsiumin signaloinnin ratkaisevaa osuutta, tutkittiin 40 eri yhdisteen aiheuttamaa apoptoosia ja nämä aineet ovat seulottavissa esiin kemiallisesta kirjastosta. Näistä tutkimuksista voidaan päätellä, että KALPAIINI ( calpain) ja KALMODULIINIKINAASI II (CaMKII) ovat tärkeitä tekijöitä kalsiumin signaloinnin välittämisessä apoptoositapahtumassa.
• Kalsiumin osuus apoptoottisessa prosessissa oli myöhäinen ja kalsiumin signalointi havaittiin taustatekijänä Jun N-terminaalisen kinaasin (JNK) aktivaatiossa.
http://journals.prous.com/journals/dnp/20041710/html/dn170646/images/Li_f1.gif
{kind=link}
(KOMMENTTI: PI3K sanotaan survival tekijäksi. Sen sijaan IP6K2 on apoptoosin tekijä, mikä on uudempaa tietoa) http://ccforum.com/content/figures/cc2387-1-l.jpg
{kind=link}
Toinen väitöskirjatyön päämäärä oli kehittää monitorointimetodia in vivo-tarkoituksiin syöpäterapialääkkeitten aktiivisuuden mittauksia varten.
KASPAASIEN OSUUS apoptoosiaktiviteetin toimittajana. (Tästä kirjoitan erikseen)
Tutkijat mittasivat kaspaasien (CASPASE) pilkkomien SYTOKERATIINI 18-fragmenttien (CK18) määrää potilaan seerumista syöpäterapian aikana. Metodista kehkeytyi kiinnostavaa kliinistä informaatiota.
Syöpälääke DOCETAXEL(x) (http://en.wikipedia.org/wiki/Docetaxel) havaittiin tehokkaammaksi tuumorin apoptoosin indusoijaksi prostatasyövässä verrattuna kahteen muuhun tutkittuun valmisteesen. Eräs erityinen mielenkiintoinen havainto oli se, että apoptoosi indusoitui esiin useitten hoitojaksojen kuluessa.
Kun mitattiin kaspaasilla esiin pilkkoutuneitten totaalisytokeratiini-18 fraktion määriä seerumista, saatiin osoitusta siitä, että
CEF- kombinaatio, jota käytettiin rintasyövän hoitoon, indusoi pääsiassa nekroottisen solutuhovasteen.(CEF= syklofosfamidi, epirubisiini, 5-fluorourasiili). http://www.germanbreastgroup.de/nnbc3/english.html
Kun parantuu metodit, joilla määritetään lääkevaikutuksia in vivo olosuhteissa on odotettavissa uusien lääkkeiden kliiniseen kehityksen apua ja samoin hyötyä kliinisen työhön.
(x) Docetaxel on semisynteettinen taksaani, kemoterapeuttinen aine, joka sitoo beeta-tubuliinia, stabiloi mikrotubuluksia ja aiheuttaa solusyklin pysähtymää (CELL CYCLE ARREST) ja apoptoosia (2001). Jos tällä käsiteltiin 3 viikon välein, tulokset hengissäpysymisen ja paranemiseen suhteen paremmat kuin mitoxantronilla (2004).
(Kommentti: Väitöskirjassa kuvattuja lääkkeitä ovat myös
Estramustiini polyfosfaatti (EMP) on (nornitrogen mustard)-estradiolikonjugaatti, joka vaikuttaa mikrotubulaariseen dynamiikkaan ( 1999). Sitä on käytetty prostatasyöpään vuosia.
http://en.wikipedia.org/wiki/Estramustine
Vinorelbine vaikuttaa myös mikrotubuluksiin ja sillä on aktiivisuutta prostatasyöpää vastaan.
http://en.wikipedia.org/wiki/Vinorelbine
Väitöskirjatyössä verrattiin estramustiinia, vinorelbiinia ja docetakselia).
VAIKUTUSMEKANISMIN TAUSTASTA KALSIUMIN SUHTEEN
( On paljon strategioita, joita koetetaan kayttää, mutta monsisa konvergoituu viime käden tärkeäksi signaloivaksi vaikuttajaksi Calsium joni ja täten inositolifosfaatti järjestelmän molekylääriset resurssit).
http://www.nature.com/cdd/journal/v13/n8/fig_tab/4401940f3.html#figure-title)
Väitöskirjatyöhön liittyy kuva KALSIUMIN SIGNALOINNISTA TÄSSÄ YHTEYDESSÄ. Siihen kuuluvia osatekijöitä ovat mm:
CALPAIN , Ca++aktivoituva proteaasi kalpaiini on tärkeä apoptoosin signaalivälittäjä.
http://nashua.case.edu/pathwaysweb/GOPathwaysAnalysisTool/resource/GO/images/0004198.png
{kind=link}
Kalpaiini vaikuttaa eräänlaiselta solun serubbaabelilta, (ett slags härjande på fältet).
www.nature.com/.../v28/n4/fig_tab/9600595f2.html
CALPEPTIN, kalpeptiini on kalpaiinin inhibiittori ja voi estää 19 yhdistettä niistä 40 tutkitusta apoptoosia aiheuttavasta aineesta.
Kalpaiini ei yksinään kuitenkaan voi vastata kaikista apoptoosivaikutuksista.
CALMODULIN (CaM), kalmoduliini on tärkeä ja pääasiallinen intrasellulaarinen Ca++-sitova proteiini ja se voi olla agonisti tai antagonisti apoptoosille.
Jos W7- tekijällä estettiin kalmoduliini, apoptoosia aiheuttavista 40 tekijästä estyi 17 tekijää. Kalmoduliinin osuus kuitenkin oli tärkeä apoptoosissa.
http://www.nature.com/nrn/journal/v6/n4/images/nrn1647-f2.jpg
{kind=link}
CALCINEURIN (proteiinifosfataasi 2B, PP2B), kalneuriini. Tämä on Ca-kalmoduliinilla aktivoituva seriini-treoniini PP ja aktivoituu solunsisäisestä kalsiumista (Ca++)i. Kalkineuriini poistaa fosfaattia ASK-1:stä, joka on apoptoosia signaloiva kinaasi-1 ja tällöin tämän kinaasin aktiviteetti pääsee lisääntymään.
ASK1 sitten aktivoi JNK ja p38 ja johtaa kohti apoptoosia.
Kalkineuriinia pidetään solukuoleman moduloijana. Se on myös sensitiivinen solustressille, vaikka tässä tapauksesa oksidaatio johtaa entsyymin inaktivaatioon. Mitään vahvaa apoptoosin estymistä ei siitä kuitenkaan saa aikaan inhibiittorilla.
(Kommentti: Stressiproteiineista se voi aktivoitua ja johtaa tällöin solun selviämiseen.
(http://molinterv.aspetjournals.org/cgi/content/full/2/5/284/F1
“The unfolded protein response (UPR) is a transcriptional and translational – tämä tekijä on otettava huomoon, sillä nykyajan ihmisellä on aina sen verran solustressejä- mikä sitten reaktioketjun loppusuoralla vaikuttaa solun apoptoosin ja eloonjäämisen exekuutiossa- asian voi miltei pelkistää IP3/ IP7 suhteeksi - ehkä myös sanoilla, kalsiumin säädelty vaimennus / kalsiumin progrediointiin sysätty runsas lisäys ).
USEITA KALMODULIINISTA riippuvia proteiineja on identifioitu ja yksi näistä on
CALMODULIN KINASE II ( CaMK II), joka toimii välittäjänä solun kalsiumin ja kalmoduliinivälitteisen apoptoosin kesken. CaMK II voi fosforyloida ASK-1 kinaasia, mutta suhde apoptoosiin ei ole tutkittu.
Kuitenkin CaMK II inhibiittori (KN-93) kykeni blokeeraamaan apoptoosin 21 tapauksessa 40 :stä.
JNK-välitteiselle apoptoosille löytyy useita signaalireittejä, kuten p53 ( ns. genomin suojelijamolekyyli) sekä Bak ja Bax.
http://genesdev.cshlp.org/content/13/8/916/F1.large.jpg
http://www-rcf.usc.edu/~forsburg/images/Image10.jpg
{kind=link}
http://herkules.oulu.fi/isbn9514270398/html/graphic22.png
{kind=link}
http://www.nature.com/bjc/journal/v94/n4/images/6602955f7.gif
{kind=link}
JNK-välitteisen apopoosin pystyi estämään eräät aineet:kuten JNK-estäjä SP600125.
Kts HSP stressiproteiinien osuutta:
http://www.biolsci.org/v03/p0020/ijbsv03p0020g01.jpg
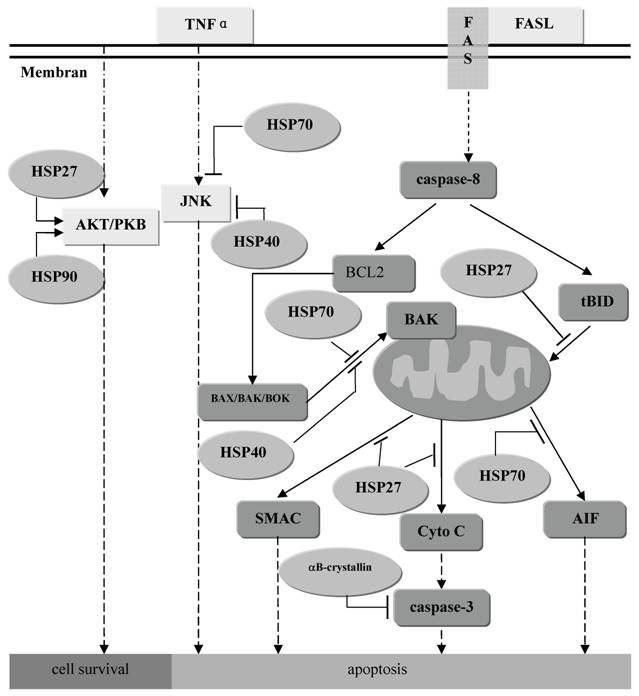{kind=link}
CALSIUMIN SIGNALOINTI aktivoi JNK:n - joissain tutkimuksissa. Osa signaaleista kulkee CaMK II / JNK tietä.Tutkija mainitsi, että ollaan tekemässä lisätutkimuksia, joissa selvitellään, onko JNK aktivaation edellä CaMKII vaikutus.
5.6.2009 11:18
torsdag 4 juni 2009
Solun apoptoosin säätely IP7 ja IP6K2 avulla
Solun elossapysymisen säätely HSP90 ja IP6K2:n avulla.
Apoptoosin säätelystä ja lääkevaikutuksesta
" the pyrophosphate diphosphoinositol pentakisphosphate (5-PP-IP5, IP7), .... Novobiocin increases nuclear and decreases cytosolic IP6K2"
http://www.wipo.int/pctdb/en/wo.jsp?IA=US2007025246&WO=2008073382&DISPLAY=DESC
Tätä yllä mainittua työtä kannatetaan US kansanterveyspalvelujen ja tieteelliseen tutkimukseen annetulla apurahalla . USA:n hallitus pidättää tietyt oikeudet inventioon. Invention teknisestä puolesta: Inventio käsittää proteiini-proteiini-interaktioita, erityisesti syöpälääkkeitten seulontaa. Suomennan alusta muutaman kappaleen.
Invention tausta:
INOSITOLIFOSFAATIT ovat tärkeitä solunsisäisiä signaalimolekyylejä ja näistä on kaikkein tunnetuin inositoli-1,4,5-trifosfaatti(IP3) , joka pystyy vapauttamaan solunsisäistä kalsiumia (Ca++). Insitolifosfaattien joukossa on viime aikoina alettu fokusoitua korkeampiin polyfosfaatteihin sekä inositolipyrofosfaattiin (IP7). Tämä IP7 on toiselta nimeltä 5-PP-IP5 (pyrophosphate diphosphoinositoli pentakisphosphate). Tämä muoto pystyy antamaan energeettisen fosfaatin eri proteiinikohteille.
IP7 muodostuu kehossa kolmella IP6K kinaasiperheen entsyymillä (IP6K1, IP6K2, IP6K3),
joista IP6K2, näyttää olevan assosioitunut apoptoosiin.
Apoptoottiset stimulukset huomattavasti lisäävät IP7-molekyylin (5-PP-IP5) muodostumista Entsyymin IP6K2 yliexpressio lisää solukuolemaa ja siRNA-indusoima depleetio taas edistää solun hengissäpysymistä.
Ns. lämpöshokkiproteiinit HSP, stressiproteiinit, omaavat klassisen roolin denaturoitujen proteiinien uudelleen laskostumisen edistäjinä. Tämän roolin lisäksi niillä on osoittautunut olevan osaa antiapoptoottisessa kaskadissa ja niistä on tullut anti-syöpälääkkeiden kehittelyssä kohteita.
http://en.wikipedia.org/wiki/Heat_shock_protein
On jatkuvasti tarvetta löytää uusia lääkeaineita syövän hoitoon.
YHTEENVETO INVENTIOSTA
(1) Yksi inventiomalli edellyttää metodia, jolla voidaan identifioida yhdisteitä, jotka vuorovaikuttavat IP6K2-proteiinin sitoutumisessa lämpöshokkiproteiiniin HSP90. Sellaiset yhdisteet ovat terapeuttisia kandidaattiaineita.
Ykkösproteiini, kakkosproteiini ja testiyhdiste asetetaan kontaktiin toistensa kanssa sillä edellytyksellä, että ykkös- ja kakkosyhdiste sitoutuvat toisiinsa, jos testiaine ei ole läsnä. ykkös ja kakkosproteiineina toimivat IP6K2 ja HSP90.
Määritetään ykkösproteiinista määrät, jotka sitoutuvat , tai irtoavat tai estyvät sitoutumasta kakkosproteiiniin. Terapeuttinen kandidaattiaine identifioidaan, jos yhdiste vähentää ykkösproteiinin sitoutumista kakkosproteiiniin tai vaikuttaa ykkösproteiinin irtoamisen kakkosproteiinsita tai estää ykkösproteiinin sitoutumasta kakkosproteiiniin.
ESIM.1— HSP90 ja IP6K2 saostuvat yhdessä
Tutkijat pohtivat, jos IP6K2 entsyymin nopea aktivaatio vasteena apoptoottiselle stimulukselle heijastaisi interaktiota toisiin proteiineihin. He kehittivät GDDA:n (Gestalt Domain Detection Algorithm) joka nopeutti havaitsemaan proteiini-proteiini tunnistusmotiiveja. Tällä GDDA-analyysillä havaittiin IP6K2:n aminohapposekvenssi 131-140, mikä muistutti hyvin konservoitua motiivia p23:ssa, joka on HSP90 chaperoniin korkealla affiniteetilla sitoutuva co-chaperoni.
Tämä sekvenssi on hyvin samanlainen IP6K2 ja IP6K3 entsyymeillä, mutta ei IP6K1 entsyymillä. Sekvenssianalyysin kanssa yhtenevänä tietona saatiin myös, että endogeeninen HSP90 saostuu yhdessä Myc-IP6K2 ja IP6K3 kanssa, mutta ei IP6K1:n kanssa.
Endogeeninen HSP90 HeLa-soluissa sitoutuu yliexpressoituvaan myc-IP6K2:teen .
Endogeeninen HSP90 ja endogeeninen IP6K2 saostuvat myös yhdessä hiiren aivosta.
Nisäkäsperäinen HSP90:n sitoutuminen immuunisaostettuun Myc-IP6K2:een vahvistaa, että interaktio on suora.
(2) Eräs inventiometodi on vastaava, mutta proteiinina on polypeptidejä ykkös- ja kakkos polypeptidit ovat kooltaan vähempiä kuin koko HSP90 tai koko IP6K2 proteiinit, mutta kuitenkin riittävän pitkiä sitoutuakseen.
ESIM.2. HSP90 estää IP6K2:n katalyyttisen aktiivisuuden
Jotta tutkijat saisivat varmuuden siitä, että HSP90 sitoutuminen IP6K2:teen säätelee IP6K2 aktiivisuutta myös in vivo ( kehossa), he merkkasivat HeLA ja HEK293 soluja tritium-inositolilla ja monitoroivat IP6 molekyylin muuttumisen IP7 molekyyliksi.( PP—IP5 muotoon) . Molemmissa solulinjoissa HSP90-yliexpressio vähensi noin puoleen IP6K2-entsyymin aktiivisuutta transfektoimattomissa soluissa ja myös soluissa, joihin oli transfektoitu IP6K2 .
Päinvastoin RNA-interferenssillä tehty HSP90-vaje lisäsi IP6K-aktiivisuutta.
Nämä löydöt viittaavat siihen, että HSP90:n sitoutuminen IP6K2 entsyymiin fysiologisesti estää tämän entsyymin katalyyttisen aktiivisuuden. Tutkijat ottivat selvää siitä mahdollisuudesta, jos IP6K2 aktiivisuus on hiivasolussa samalla tavalla säätynyt. Hiivasolu HSC82 muistuttaa ihmisen HSP90 sekä aminohappojärjestykseltään että läsnäololtaan basaaliolosuhteiden vallitessa. HSC82:n deleetio kohottaa IP7 molekyylin muodostumisen kaksinkertaiseksi, kun taas asiaan liittymättömän HSP:n (HSP1 04) deleetio ei lisää, vaan sensijaan hieman vähentää IP7:n muodostumsita.
(3) Eräs inventioaspekti käsitti metodin, jolla identifioitiin yhdisteitä, jotka vuorovaikuttavat humaanin IP6K2 entsyymiproteiinin ja humaani HSP90-proteiinin sitoutumiseen.
Testiyhdiste johdetaan solun kanssa kontaktiin. Solu käsittää kolme rekombinantti DNA-kosntruktia. Ensimmäinen konstrukti koodaa ykköspolypeptidiä, joka on fusoitu sekvenssispesifiseen DNA:ta sitovaan domaaniin . Toinen konstrukti koodaa kakkospolypeptidiä, joka on fusoitu transkription aktivoivaan domaaniin. Kolmas konstrukti käsittää reportterigeenin, joka sijaitsee alavirtaan DNA-elementistä, joka tunnistuu sekvenssispesifisellä DNA:ta sitovalla domaanilla. Ykköspolypeptidi on IP6K2 ja kakkospolypeptidi on HSP90 tai päinvastoin tai niistä riittävän pitkä polypeptidi. Määritetään reportterigeenin määrä yhdisteessä.
ESIM.3. Identifioidaan IP6K2:n sitova domaani
Tutkijat kartoitttivat IP6K2:n HSP90:tä sitovan domaanin ja vertasivat mutaatioentsyymiin. Jos oli tryptofaani-131 (W131) , IP6K2-HSP90 sitoutuminen oli kohtalaisesti vähentynyt.
Jos oli arginiini-133 (R133) , arginiini-136(R136) tai glutamaatti 138, 139(E138,139), sitoutumiskyky poistui.
Mutaatiossa IP6K2 W131A oli entsyymin IP6K2 katalyyttinen aktiivisuus vahvasti vähentynyt, mikä heijastuu sen vähentyneenä sitoutumisena substraattiinsa fytiiniin, IP6.
Mutaatiossa IP6K2 R133A ja IP6K2 E138-9 A on sama katalyyttinen aktiivisuus ja IP6:ta sitova kyky kuin IP6K2wt luonnollisessa entsyymissä.
(4) Eräs inventioaspekti on tämäkin: Määritetään ihmisen HSP90-proteiinin kvantitatiivinen määrä, jolla se sitoutuu ihmisen IP6K2-proteiiniin tai päinvastoin . Ykkösproteiini viedään kontaktiin kakkosproteiinin kanssa. Määritetään ykkösproteiinin se määrä, mikä sitoutuu kakkosproteiiniin.
ESIM 4— HSP90 sitoutuminen IP6K2-proteiiniin kumoaa solukuoleman(solun apoptoosin).
Ensin tutkijat ottivat selvää IP6K2 W131A mutanttientsyymin aktiivisuudesta koeputkitasossa. Yhtäpitävästi katalyyttisen aktiviteetin vähenemän kanssa in vitro, vastaavaa mutanttia transfektoitaessa saatiin paljon vähemmän entsyymiaktiivisuutta kuin luonnollisella entsyymillä IP6K2wt.
Ja päinvastoin, jos solut ilmensivät ylimäärin IP6K2-R133A tai IP6K2-E138-9A,jotka eivät pysty sitomaan HSP90, oli entsyymiaktiivisuus soluissa miltei kaksinkertainen verrattuna luonnolliseen IP6K2 entsyymiin.
Kun HSP:n yliesiintymä huomattavasti vähentää IP7 molekyylin muodostumista soluissa luonnolliseslla IP6K2-entsyymillä, niin mitään alenemaa IP7-muodostuksessa ei tapahtunut niissä soluissa, joissa esiintyi ylimäärin Rl33 A or E138-9A mutaatiomuotoja.
Täten havaitaan, että HSP90 (stressiproteiinin) sitoutuminen IP6K2:een fysiologisesti säätelee matalaksi IP7 molekyylin muodostumista.
Jotta voitiin määritellä, sääteleekö HSP90-IP6K2 sitoutuminen IP7-assosioitunutta solukuolemaa, transfektoitiin HeLA-soluja IP6K2 wt tai mutanttientsyymimuodoilla.
Jos luonnollista IP6K2wt entsyymiä yliexpressoitui, solukuolema kumoutui, ja tämän vaikutuksen muutti HSP90, jos sitä esiintyi saman aikaan.
Hämmästyttävästi se solukuolema, joka seuraa IP6K2 entsyymin mutanttimuotojen R133A and E138-9A yliexpressoitumisesta, oli viisi kertaa suurempi, mitä luonnollisen IP6K2wt entsyymin yliekspressiosta seurasi, ja niissä soluissa HSP90 ei onnistunut vähentämään solukuolemaa.
Niinpä voitiin todeta, että juuri endogeenisen stressiproteiinin HSP90 kyky sitoutua entsyymiin IP6K2, poistaa fysiologisesti solun apoptoottiset aktiot ( ja IP7 muodostus vähenee) .
Tätä johtopäätöstä tukee tieto siitä, että HSP90:n yli-expressoituminen ei onnistu vähentämään sellaista solukuolemaa, joka aiheutuu R 133 A and E138-9A mutanteista , jotka eivät voi sitoa HSP90:tä.
IP6K2 entsyymin katalyyttisen toiminnan tärkeys solukuolemassa, apoptoosissa, huomataan siitäkin, kun solukuoleman lisääntyminen jää tapahtumatta, jos katalyyttisesti vajeiset mutantit IP6K2 Wl 3 IA yliexpressoituvat.
(5) Eräs tutkimus tehtiin siten, että oli fuusioproteiini ykkös ja kakkospolypeptidistä. Kakkososana oli nisäkäsperäinen IP6K2, joka pystyy sitomaan HSP:tä ja ykköspolypeptidinä ei ollut osia IP6K2 proteiinista.
ESIM 5. Lääkkeen apoptoottinen potentiaali korreloi sen kykyyn estää IP6K2-HSP90 sitoutumista.
Koko joukko apoptooseja indusoivista anti-syöpälääkkeistä vaikuttavat ainakin osittain sitoutumalla HSP90- tekijään. Sellaisia lääkkeitä ovat geldanamycin johdannainen AAG (17-aminoallyl geldanamycin), novobiocin, ja cisplatin.
Tutkijat pohtivat, jos näitten ja muitten lääkkeiden apoptoottiset aktiot käsittävät puuttumisen HSP90-IP6K2 interaktioon.
HEK293 soluilla tehdyissä immuunisaostuskokeissa vähensivät cisplatin, novobiocin ja staurosporin IP6K2-HSP90- sitoutumista, kun taas AAG lisäsi tätä sitoutumista. Tutkijat selvittivät, että lääkkeet suoraan blokeerasivat IP6K2-HSP90 ja he mittasivat niiden suhteelliset tehokkuudet käyttämällä puhdistettuja proteiineja.
Novobiocin, cisplatin ja staurosporine vähensivät sitoutumista 50% (vastaavissa) pitoisuuksissaan (200 μM, 1- 5 μM ja 0.1 μM ).
Nämä tehokkuudet vastaavat hyvin näiden lääkkeiden soluviljelmissä osoittamiin tehoihin.
AAG ei estänyt sitoutumista intakteissa soluissa eikä koeputkessa.
Aiemmin ei ole raportoitu staurosporiinin sitoutuvan stressiproteiiniin HSP90. Tutkittiin sen vaikutus HSP90-sitoutumisees ATP-agaroosiin ja havaittiin 50% vähenemä sitoutumisessa staurosporiinin 0.2 μM pitoisuudessa , samaan tapaan kuten lääke oli osoittanut tehokkuuttaan myös irrottamalla IP6K2-HSP90- sitoutumistakin , mikä on johdonmukaista sen sitoutuessa suoraan HSP90-molekyyliin . Repeytyminen irti HSP90- sidoksesta vaikuttaa sykäyksen IP6K2-aktiviteettiin. Täten HeLa soluissa ( joissa on tai ei ole IP6K2 yliexpressio) cisplatiini, novobiosiini ja staurosporiini saavat aikaan lisääntyneen IP6K2 aktiivisuuden, kun taas AAG, joka lisää IP6K2-HSP90-sitoutumista, assosioituu vähentyneeseen IP7-muodostukseen
IP6K2-HSP90 sitoutumisen blokeeraaminen lääkkeillä oletettavasti johtaa lääkkeiden vaikuttamaan IP7 molekyylin generoitumisen lisääntymiseen. HEK-soluista on saatu samanlaisia tuloksia.
Apoptoosin säätelystä ja lääkevaikutuksesta
" the pyrophosphate diphosphoinositol pentakisphosphate (5-PP-IP5, IP7), .... Novobiocin increases nuclear and decreases cytosolic IP6K2"
http://www.wipo.int/pctdb/en/wo.jsp?IA=US2007025246&WO=2008073382&DISPLAY=DESC
Tätä yllä mainittua työtä kannatetaan US kansanterveyspalvelujen ja tieteelliseen tutkimukseen annetulla apurahalla . USA:n hallitus pidättää tietyt oikeudet inventioon. Invention teknisestä puolesta: Inventio käsittää proteiini-proteiini-interaktioita, erityisesti syöpälääkkeitten seulontaa. Suomennan alusta muutaman kappaleen.
Invention tausta:
INOSITOLIFOSFAATIT ovat tärkeitä solunsisäisiä signaalimolekyylejä ja näistä on kaikkein tunnetuin inositoli-1,4,5-trifosfaatti(IP3) , joka pystyy vapauttamaan solunsisäistä kalsiumia (Ca++). Insitolifosfaattien joukossa on viime aikoina alettu fokusoitua korkeampiin polyfosfaatteihin sekä inositolipyrofosfaattiin (IP7). Tämä IP7 on toiselta nimeltä 5-PP-IP5 (pyrophosphate diphosphoinositoli pentakisphosphate). Tämä muoto pystyy antamaan energeettisen fosfaatin eri proteiinikohteille.
IP7 muodostuu kehossa kolmella IP6K kinaasiperheen entsyymillä (IP6K1, IP6K2, IP6K3),
joista IP6K2, näyttää olevan assosioitunut apoptoosiin.
Apoptoottiset stimulukset huomattavasti lisäävät IP7-molekyylin (5-PP-IP5) muodostumista Entsyymin IP6K2 yliexpressio lisää solukuolemaa ja siRNA-indusoima depleetio taas edistää solun hengissäpysymistä.
Ns. lämpöshokkiproteiinit HSP, stressiproteiinit, omaavat klassisen roolin denaturoitujen proteiinien uudelleen laskostumisen edistäjinä. Tämän roolin lisäksi niillä on osoittautunut olevan osaa antiapoptoottisessa kaskadissa ja niistä on tullut anti-syöpälääkkeiden kehittelyssä kohteita.
http://en.wikipedia.org/wiki/Heat_shock_protein
On jatkuvasti tarvetta löytää uusia lääkeaineita syövän hoitoon.
YHTEENVETO INVENTIOSTA
(1) Yksi inventiomalli edellyttää metodia, jolla voidaan identifioida yhdisteitä, jotka vuorovaikuttavat IP6K2-proteiinin sitoutumisessa lämpöshokkiproteiiniin HSP90. Sellaiset yhdisteet ovat terapeuttisia kandidaattiaineita.
Ykkösproteiini, kakkosproteiini ja testiyhdiste asetetaan kontaktiin toistensa kanssa sillä edellytyksellä, että ykkös- ja kakkosyhdiste sitoutuvat toisiinsa, jos testiaine ei ole läsnä. ykkös ja kakkosproteiineina toimivat IP6K2 ja HSP90.
Määritetään ykkösproteiinista määrät, jotka sitoutuvat , tai irtoavat tai estyvät sitoutumasta kakkosproteiiniin. Terapeuttinen kandidaattiaine identifioidaan, jos yhdiste vähentää ykkösproteiinin sitoutumista kakkosproteiiniin tai vaikuttaa ykkösproteiinin irtoamisen kakkosproteiinsita tai estää ykkösproteiinin sitoutumasta kakkosproteiiniin.
ESIM.1— HSP90 ja IP6K2 saostuvat yhdessä
Tutkijat pohtivat, jos IP6K2 entsyymin nopea aktivaatio vasteena apoptoottiselle stimulukselle heijastaisi interaktiota toisiin proteiineihin. He kehittivät GDDA:n (Gestalt Domain Detection Algorithm) joka nopeutti havaitsemaan proteiini-proteiini tunnistusmotiiveja. Tällä GDDA-analyysillä havaittiin IP6K2:n aminohapposekvenssi 131-140, mikä muistutti hyvin konservoitua motiivia p23:ssa, joka on HSP90 chaperoniin korkealla affiniteetilla sitoutuva co-chaperoni.
Tämä sekvenssi on hyvin samanlainen IP6K2 ja IP6K3 entsyymeillä, mutta ei IP6K1 entsyymillä. Sekvenssianalyysin kanssa yhtenevänä tietona saatiin myös, että endogeeninen HSP90 saostuu yhdessä Myc-IP6K2 ja IP6K3 kanssa, mutta ei IP6K1:n kanssa.
Endogeeninen HSP90 HeLa-soluissa sitoutuu yliexpressoituvaan myc-IP6K2:teen .
Endogeeninen HSP90 ja endogeeninen IP6K2 saostuvat myös yhdessä hiiren aivosta.
Nisäkäsperäinen HSP90:n sitoutuminen immuunisaostettuun Myc-IP6K2:een vahvistaa, että interaktio on suora.
(2) Eräs inventiometodi on vastaava, mutta proteiinina on polypeptidejä ykkös- ja kakkos polypeptidit ovat kooltaan vähempiä kuin koko HSP90 tai koko IP6K2 proteiinit, mutta kuitenkin riittävän pitkiä sitoutuakseen.
ESIM.2. HSP90 estää IP6K2:n katalyyttisen aktiivisuuden
Jotta tutkijat saisivat varmuuden siitä, että HSP90 sitoutuminen IP6K2:teen säätelee IP6K2 aktiivisuutta myös in vivo ( kehossa), he merkkasivat HeLA ja HEK293 soluja tritium-inositolilla ja monitoroivat IP6 molekyylin muuttumisen IP7 molekyyliksi.( PP—IP5 muotoon) . Molemmissa solulinjoissa HSP90-yliexpressio vähensi noin puoleen IP6K2-entsyymin aktiivisuutta transfektoimattomissa soluissa ja myös soluissa, joihin oli transfektoitu IP6K2 .
Päinvastoin RNA-interferenssillä tehty HSP90-vaje lisäsi IP6K-aktiivisuutta.
Nämä löydöt viittaavat siihen, että HSP90:n sitoutuminen IP6K2 entsyymiin fysiologisesti estää tämän entsyymin katalyyttisen aktiivisuuden. Tutkijat ottivat selvää siitä mahdollisuudesta, jos IP6K2 aktiivisuus on hiivasolussa samalla tavalla säätynyt. Hiivasolu HSC82 muistuttaa ihmisen HSP90 sekä aminohappojärjestykseltään että läsnäololtaan basaaliolosuhteiden vallitessa. HSC82:n deleetio kohottaa IP7 molekyylin muodostumisen kaksinkertaiseksi, kun taas asiaan liittymättömän HSP:n (HSP1 04) deleetio ei lisää, vaan sensijaan hieman vähentää IP7:n muodostumsita.
(3) Eräs inventioaspekti käsitti metodin, jolla identifioitiin yhdisteitä, jotka vuorovaikuttavat humaanin IP6K2 entsyymiproteiinin ja humaani HSP90-proteiinin sitoutumiseen.
Testiyhdiste johdetaan solun kanssa kontaktiin. Solu käsittää kolme rekombinantti DNA-kosntruktia. Ensimmäinen konstrukti koodaa ykköspolypeptidiä, joka on fusoitu sekvenssispesifiseen DNA:ta sitovaan domaaniin . Toinen konstrukti koodaa kakkospolypeptidiä, joka on fusoitu transkription aktivoivaan domaaniin. Kolmas konstrukti käsittää reportterigeenin, joka sijaitsee alavirtaan DNA-elementistä, joka tunnistuu sekvenssispesifisellä DNA:ta sitovalla domaanilla. Ykköspolypeptidi on IP6K2 ja kakkospolypeptidi on HSP90 tai päinvastoin tai niistä riittävän pitkä polypeptidi. Määritetään reportterigeenin määrä yhdisteessä.
ESIM.3. Identifioidaan IP6K2:n sitova domaani
Tutkijat kartoitttivat IP6K2:n HSP90:tä sitovan domaanin ja vertasivat mutaatioentsyymiin. Jos oli tryptofaani-131 (W131) , IP6K2-HSP90 sitoutuminen oli kohtalaisesti vähentynyt.
Jos oli arginiini-133 (R133) , arginiini-136(R136) tai glutamaatti 138, 139(E138,139), sitoutumiskyky poistui.
Mutaatiossa IP6K2 W131A oli entsyymin IP6K2 katalyyttinen aktiivisuus vahvasti vähentynyt, mikä heijastuu sen vähentyneenä sitoutumisena substraattiinsa fytiiniin, IP6.
Mutaatiossa IP6K2 R133A ja IP6K2 E138-9 A on sama katalyyttinen aktiivisuus ja IP6:ta sitova kyky kuin IP6K2wt luonnollisessa entsyymissä.
(4) Eräs inventioaspekti on tämäkin: Määritetään ihmisen HSP90-proteiinin kvantitatiivinen määrä, jolla se sitoutuu ihmisen IP6K2-proteiiniin tai päinvastoin . Ykkösproteiini viedään kontaktiin kakkosproteiinin kanssa. Määritetään ykkösproteiinin se määrä, mikä sitoutuu kakkosproteiiniin.
ESIM 4— HSP90 sitoutuminen IP6K2-proteiiniin kumoaa solukuoleman(solun apoptoosin).
Ensin tutkijat ottivat selvää IP6K2 W131A mutanttientsyymin aktiivisuudesta koeputkitasossa. Yhtäpitävästi katalyyttisen aktiviteetin vähenemän kanssa in vitro, vastaavaa mutanttia transfektoitaessa saatiin paljon vähemmän entsyymiaktiivisuutta kuin luonnollisella entsyymillä IP6K2wt.
Ja päinvastoin, jos solut ilmensivät ylimäärin IP6K2-R133A tai IP6K2-E138-9A,jotka eivät pysty sitomaan HSP90, oli entsyymiaktiivisuus soluissa miltei kaksinkertainen verrattuna luonnolliseen IP6K2 entsyymiin.
Kun HSP:n yliesiintymä huomattavasti vähentää IP7 molekyylin muodostumista soluissa luonnolliseslla IP6K2-entsyymillä, niin mitään alenemaa IP7-muodostuksessa ei tapahtunut niissä soluissa, joissa esiintyi ylimäärin Rl33 A or E138-9A mutaatiomuotoja.
Täten havaitaan, että HSP90 (stressiproteiinin) sitoutuminen IP6K2:een fysiologisesti säätelee matalaksi IP7 molekyylin muodostumista.
Jotta voitiin määritellä, sääteleekö HSP90-IP6K2 sitoutuminen IP7-assosioitunutta solukuolemaa, transfektoitiin HeLA-soluja IP6K2 wt tai mutanttientsyymimuodoilla.
Jos luonnollista IP6K2wt entsyymiä yliexpressoitui, solukuolema kumoutui, ja tämän vaikutuksen muutti HSP90, jos sitä esiintyi saman aikaan.
Hämmästyttävästi se solukuolema, joka seuraa IP6K2 entsyymin mutanttimuotojen R133A and E138-9A yliexpressoitumisesta, oli viisi kertaa suurempi, mitä luonnollisen IP6K2wt entsyymin yliekspressiosta seurasi, ja niissä soluissa HSP90 ei onnistunut vähentämään solukuolemaa.
Niinpä voitiin todeta, että juuri endogeenisen stressiproteiinin HSP90 kyky sitoutua entsyymiin IP6K2, poistaa fysiologisesti solun apoptoottiset aktiot ( ja IP7 muodostus vähenee) .
Tätä johtopäätöstä tukee tieto siitä, että HSP90:n yli-expressoituminen ei onnistu vähentämään sellaista solukuolemaa, joka aiheutuu R 133 A and E138-9A mutanteista , jotka eivät voi sitoa HSP90:tä.
IP6K2 entsyymin katalyyttisen toiminnan tärkeys solukuolemassa, apoptoosissa, huomataan siitäkin, kun solukuoleman lisääntyminen jää tapahtumatta, jos katalyyttisesti vajeiset mutantit IP6K2 Wl 3 IA yliexpressoituvat.
(5) Eräs tutkimus tehtiin siten, että oli fuusioproteiini ykkös ja kakkospolypeptidistä. Kakkososana oli nisäkäsperäinen IP6K2, joka pystyy sitomaan HSP:tä ja ykköspolypeptidinä ei ollut osia IP6K2 proteiinista.
ESIM 5. Lääkkeen apoptoottinen potentiaali korreloi sen kykyyn estää IP6K2-HSP90 sitoutumista.
Koko joukko apoptooseja indusoivista anti-syöpälääkkeistä vaikuttavat ainakin osittain sitoutumalla HSP90- tekijään. Sellaisia lääkkeitä ovat geldanamycin johdannainen AAG (17-aminoallyl geldanamycin), novobiocin, ja cisplatin.
Tutkijat pohtivat, jos näitten ja muitten lääkkeiden apoptoottiset aktiot käsittävät puuttumisen HSP90-IP6K2 interaktioon.
HEK293 soluilla tehdyissä immuunisaostuskokeissa vähensivät cisplatin, novobiocin ja staurosporin IP6K2-HSP90- sitoutumista, kun taas AAG lisäsi tätä sitoutumista. Tutkijat selvittivät, että lääkkeet suoraan blokeerasivat IP6K2-HSP90 ja he mittasivat niiden suhteelliset tehokkuudet käyttämällä puhdistettuja proteiineja.
Novobiocin, cisplatin ja staurosporine vähensivät sitoutumista 50% (vastaavissa) pitoisuuksissaan (200 μM, 1- 5 μM ja 0.1 μM ).
Nämä tehokkuudet vastaavat hyvin näiden lääkkeiden soluviljelmissä osoittamiin tehoihin.
AAG ei estänyt sitoutumista intakteissa soluissa eikä koeputkessa.
Aiemmin ei ole raportoitu staurosporiinin sitoutuvan stressiproteiiniin HSP90. Tutkittiin sen vaikutus HSP90-sitoutumisees ATP-agaroosiin ja havaittiin 50% vähenemä sitoutumisessa staurosporiinin 0.2 μM pitoisuudessa , samaan tapaan kuten lääke oli osoittanut tehokkuuttaan myös irrottamalla IP6K2-HSP90- sitoutumistakin , mikä on johdonmukaista sen sitoutuessa suoraan HSP90-molekyyliin . Repeytyminen irti HSP90- sidoksesta vaikuttaa sykäyksen IP6K2-aktiviteettiin. Täten HeLa soluissa ( joissa on tai ei ole IP6K2 yliexpressio) cisplatiini, novobiosiini ja staurosporiini saavat aikaan lisääntyneen IP6K2 aktiivisuuden, kun taas AAG, joka lisää IP6K2-HSP90-sitoutumista, assosioituu vähentyneeseen IP7-muodostukseen
IP6K2-HSP90 sitoutumisen blokeeraaminen lääkkeillä oletettavasti johtaa lääkkeiden vaikuttamaan IP7 molekyylin generoitumisen lisääntymiseen. HEK-soluista on saatu samanlaisia tuloksia.
IP6K2 (3p21.31) ( inositolihexakisfosfaattikinaasi 2) ja PP-InsPx , Inositolipyrofosfaatit. apoptoositekijäin säätelijänä
IP6K2 (inositolhexakisphosphatekinase 2), PiUS
On mahdollista, että inositoli 5-pyrofosfaatti (PP-IP5) voi estää DNA korjaantumisen ja siten edistää apoptoosia syövän hoidossa herkistämällä syöpäsoluja.
It is possible that InsP5PP, which contains a pyrophosphate linkage, may function as an ... InsP5PP may inhibit DNA repair and thereby promote apoptosis.
INOSITOLI hexacisfosfaattikinaasi 2 (IP62K) säätelee stimuloivasti IFN-beeta ja gammasäteilyn apoptoottista vaikutusta kaspaasi-8 tietä.
LÄHDE : Bei H Morrison,1 Joseph A Bauer et al. Inositol hexakisphosphate kinase 2 sensitizes ovarian carcinoma cells to multiple cancer therapeutics
Oncogene. 2002 March 14; 21(12): 1882–1889. Oncogene. Author manuscript; available in PMC 2007 October 29. USA.
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2043497
Tutkijat ovat identifioineet inositolihexacisfosfaattikinaasi-2:n ( IP6K2) mahdollisena apoptoosin säätelijänä.
Jos tätä entsyymiä IP6K2 ilmenee ylimäärin, lisääntyy apoptoosi, jota indusoi interferoni beeta (IFN-beeta) ja sytotoksiset agenssit ( tutkittuna ovariaalikarsinoomasoluissa).
IP6K2 tekee inositolipolyfosfaateista IP6 ja IP5 vastaavia pyrofosfaatteja.
IP6K2_HUMAN, Q9UHH9
Tämän entsyymin funktio on muuttaa IP6 muotoon ”IP7”, joka kuitenkin pitää ilmaista eri tavalla, koska inositolissa on vain 6 kohtaa, johon lisäliitos voidaan tehdä. Viidessä on yksi ja yhdessä on kaksi fosfaattia: PP-IP5 on tulos, diphosphoinositol pentakisphosphate(InsP7/PP-InsP5).
Mutta sama entsyymi voi muuttaa myös IP5 molekyylin muotoon PP-IP4, lisäämällä fosfaatin 2- asemaan inositolirengasta. (1,3,4,5,6-pentakisphosphate (InsP5) muotoon PP-InsP4)
Entsyymin muita nimiä:
ATP:1D-myo-inositol-hexakisphosphate phosphotransferase
InsP6 kinase 2
2, 3P(i)-uptake stimulator
3Pi uptake stimulator
2, inositol hexakisphosphate kinase 2
1, 2 inositol hexaphosphate kinase 2 1,
GEENI koodaa proteiinia, joka kuuluu IPK-perheeseen ( inositolifosfokinaasien perheesen) ja tekee yllä olevat muutokset IP6 ja tiettyyn IP5 molekyyliin. Se voi vaikuttaa stimuloimalla IFN-beetan kasvua suppressoivia ja apoptoottisia ominaisuuksia( Tämä on havaittu ovariaalisyövästä) Alternatiivinen pilkkoutuminen geenistä johtaa multippeleihin transkripteihin, jotka koodaavat eri isoformeja.
http://www.genecards.org/cgi-bin/carddisp.pl?gene=IP6K2
ESIMERKKI:
Tässä allaolevassa tutkimuksessa vertailtiin sekä IFN-beeta vaikutusta että gamma- säteilytystä ovariaalisyöpäsolukuoleman (apoptoosin) indusoijina ja tutkijat osoittivat että IP6K2 expressio herkisti tuumorisolut näille kummallekin vaikutukselle.
Jos säteilyttämättömiin syöpäsoluihin transfektoitiin IP6K2, muodostui vähemmän syöpäkolonioita kuin säteilyttämättömiin pelkkää vektoria ilmentäviin syöpäsoluihin
IP6K2 yliexpressoituminen aiheutti lisääntyvää radiosensitiivisyyttä ja se ilmeni alentuneena syöpäkolonioitten muodostumisena. ( CFU= colony forming units)
Sekä IFN-beeta että säteilyttäminen indusoivat kaspaasi-8 tekijää.
http://www.researchapoptosis.com/apoptosis/images/chart-ie.gif
IFN-beeta ( mutta ei gammasäteily), indusoi syöpäsoluissa TRAIL.
Gammasäteily, mutta ei IFN beeta, indusoi DR4mRNA.
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah&part=A8980&rendertype=figure&id=A8987
Ne apoptoottiset vaikutukset , mitkä tulivat IFN-beetasta ja gammasäteilystä, blokeerautuivat kun expressoitui dominantti negatiivinen mutantti DR5A reseptoria ( death receptoria 5) tai Bcl-2:ta. http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah&part=A8980&rendertype=figure&id=A8987
Kaspaasi -8 mRNA:n indusoituminen oli paljon selvempää niissä soluissa joissa ilmeni IP6K2 verrattuna pelkkää vektoria ilmentäviin soluihin.
http://www.bioscience.org/2007/v12/af/2281/fig1.jp
DISCUSSION:
In ovarian carcinoma cell lines, IFN-β and γ-irradiation induce apoptosis using similar but not identical pathways. In sensitive cells, IFN-β upregulates TRAIL, and to a lesser extent, caspase 8 (Figure 5a). IFN-β-induced apoptosis has been associated with TRAIL induction in several different tumor cell lines (Chawla-Sarkar et al., 2001; Zhang et al., 1999). γ-irradiation clearly upregulates caspase 8 and DR4 mRNA (Figure 5b) as well as caspase 8 activity (Figure 6a). This is consistent with our previous study showing induction of apoptosis following ionizing radiation (Gong and Almasan, 2000). Transcriptional induction of DR4 following radiation has also been reported in lung carcinoma cell lines (Guan et al., 2001). Our studies are consistent with others, who have described caspase 8 activation following radiation of Jurkat cells (Belka et al., 2000). In three different ovarian carcinoma cell lines, we observed no induction of TRAIL following radiation treatment. This is in contrast to hematologic cells in which both TRAIL and Fas upregulation has been described (Gong and Almasan, 2000). Combination therapy utilizing TRAIL and cytotoxic agents (cisplatin, doxorubicin, or paclitaxel) has been proposed, in an effort to overcome the chemoresistance that usually develops following treatment of women with ovarian carcinoma (Cuello et al., 2001). The p53 status of ovarian carcinoma cells did not correlate with sensitivity to death inducers. In fact, cell lines with mutant p53 (NIH-OVCAR-3, Caov-3) were more sensitive to IFN-β and γ-irradiation than cells containing wild-type p53 (Hey).
We found that DR5Δ protects against apoptosis initiated by both IFN-β and γ-irradiation, confirming that TRAIL : DR-mediated signaling occurs following treatment with both death inducers. Thus, downstream effects of TRAIL and one of its receptors can be blocked by DR5Δ. Combination treatment with both modalities resulted in additive, rather than synergistic antiproliferative effects (Figure 4b). Such additivity is consistent with upregulation of the ligand TRAIL and its receptor DR4, components of the same apoptotic pathway. DR4 and DR5, both agonistic receptors for TRAIL, form homomeric and heteromeric complexes upon ligand binding (Kischkel et al., 2000). Recent reports indicate that ligation of DR4 (Sprick et al., 2000) and DR5 (Bodmer et al., 2000) lead to caspase 8 activation, using a signaling paradigm that is similar, but not identical, to those induced by TNF-α and FasL. Caspase 8 is activated as part of the death-inducing signaling complex (DISC). We show that caspase 8 activity was induced more strongly by radiation compared to IFN-β (Figure 6a,b). Yet caspase 8 activity following combination treatment with IFN-β and γ-irradiation was no greater than that following radiation alone (not shown). It is possible that in NIH-OVCAR-3 cells, caspase 8 activity is already maximized following radiation.
Overexpression of IP6K2 enhanced the degree of apoptosis induced by several different stimuli, including chemotherapeutic drugs, IFNs, and γ-irradiation. Expression of the dominant negative SUB mutant interferes with kinase activity (Morrison et al., 2001) and abrogates death induced by all these agents. IP6K2 protein levels are elevated following IFN-β and irradiation (Figure 1). These findings suggest that IP6K2 functions as a positive regulator of cell death pathways. The mechanism by which this inositol phosphate kinase promotes apoptosis is unclear. IP6K2 converts IP6 to InsP5PP. It is possible that InsP5PP, which contains a pyrophosphate linkage, may function as an energy source that facilitates DISC assembly or signal transduction to the caspase cascade. Proteins that bind IP6 include the clathrin assembly proteins AP-2, AP-3, and AP-180 (involved in membrane fusion) (Norris et al., 1995; Voglmaier et al., 1992; Ye et al., 1995), the synaptotagmins (involved in exocytosis of synaptic vesicles) (Mehrotra et al., 2000) and arrestin (which acts as an adapter during endocytosis) (Gaidarov et al., 1999). All these proteins are closely associated with the plasma membrane. Studies in yeast have suggested that the higher inositol phosphates play a role in mRNA export from the nucleus (Saiardi et al., 2000; York et al., 1999). Finally, IP6 was found to specifically stimulate repair of double-strand breaks in DNA by the process of nonhomologous end-joining in vitro (Hanakahi et al., 2000). If the functions of InsP5PP and IP6 prove to be antagonistic, InsP5PP may inhibit DNA repair and thereby promote apoptosis.
- Also known as PIUS (Pi uptake stimulator); IHPK2; InsP6K2
- Summary: This gene encodes a protein that belongs to the inositol phosphokinase (IPK) family. This protein is likely responsible for the conversion of inositol hexakisphosphate (InsP6) to diphosphoinositol pentakisphosphate (InsP7/PP-InsP5).
- It may also convert 1,3,4,5,6-pentakisphosphate (InsP5) to PP-InsP4 and affect the growth suppressive and apoptotic activities of interferon-beta in some ovarian cancers. Alternative splicing results in multiple transcript variants encoding different isoforms. [provided by RefSeq, Jul 2008]
- Expression Ubiquitous expression in bone marrow (RPKM 16.6), skin (RPKM 16.3) and 25 other tissues See more Orthologs mouse all
- Preferred Names
- inositol hexakisphosphate kinase 2
- Names
- ATP:1D-myo-inositol-hexakisphosphate phosphotransferase
- inositol hexaphosphate kinase 2
- insp6 kinase 2
- pi uptake stimulator
ORIGIN 1 mspaframdv eprakgvlle pfvhqvgghs cvlrfnettl ckplvprehq fyetlpaemr 61 kftpqykgvv svrfeededr nlcliayplk gdhgivdivd nsdcepkskl lrwttnkkhh 121 vletektpkd wvrqhrkeek mkshkleeef ewlkksevly ytvekkgnis sqlkhynpws 181 mkchqqqlqr mkenakhrnq ykfillenlt sryevpcvld lkmgtrqhgd daseekaanq 241 irkcqqstsa vigvrvcgmq vyqagsgqlm fmnkyhgrkl svqgfkealf qffhngrylr 301 rellgpvlkk ltelkavler qesyrfysss llviydgker pevvldsdae dledlseesa 361 desagayayk pigassvdvr midfahttcr lygedtvvhe gqdagyifgl qslidivtei 421 seesge
- Conserved Domains (1) summary
-
- pfam03770
Location:203 → 418 - IPK; Inositol polyphosphate kinase
Inositol polyphosphate kinaseArgRIII has has been demonstrated to be an inositol polyphosphate kinase. (https://ec.asm.org/content/7/3/471)
- pfam03770
- Casein kinase-2 mediates cell survival through phosphorylation and degradation of inositol hexakisphosphate kinase-2. Chakraborty A, et al. Proc Natl Acad Sci U S A, 2011 Feb 8. PMID 21262846, Free PMC Article
- Inositol hexakisphosphate kinase-2, a physiologic mediator of cell death. Nagata E, et al. J Biol Chem, 2005 Jan 14. PMID 15533939
- Inositol hexakisphosphate kinase 2 sensitizes ovarian carcinoma cells to multiple cancer therapeutics. Morrison BH, et al. Oncogene, 2002 Mar 14. PMID 11896621, Free PMC Article
- Inhibition of Inositol Polyphosphate Kinases by Quercetin and Related Flavonoids: A Structure-Activity Analysis. Gu C, et al. J Med Chem, 2019 Feb 14. PMID 30624931, Free PMC Article https://www.ncbi.nlm.nih.gov/pubmed/30624931/
- IP6K structure and the molecular determinants of catalytic specificity in an inositol phosphate kinase family.
Wang H, et al. Nat Commun, 2014 Jun 24. PMID 24956979, Free PMC Article
GeneRIFs: Gene References Into Functions
- An alpha-helical pair and a rare, two-turn 310 helix, that together form a substrate-binding pocket of IP6K2.
- FGF2-signaling involves the inositol polyphosphate cascade, including inositol hexakisphosphate kinase (IP6K), and demonstrate that IP6K1,2 regulates Runx2 and osteoblast gene expression.
- Genetic polymorphisms in IP6K2 gene is associated with autoimmune disease.
- Casein kinase-2 mediates cell survival through phosphorylation and degradation of inositol hexakisphosphate kinase-2.
- Gene disruption of IP6K2 in colorectal cancer cells selectively impairs p53-mediated apoptosis, instead favoring cell-cycle arrest. IP6K2 acts by binding directly to p53 and decreasing expression of proarrest gene targets
- InsP6K2 is a physiologic mediator of cell death
- IHPK2-TRAF2 binding leads to attenuation of TAK1- and NF-kappaB-mediated signaling and is partially responsible for the apoptotic activity of IHPK2.
- IHPK2 over expression enhances sensitivity of ovarian carcinoma cells to radiation, IFN-beta, caspase 8 and DR4
On mahdollista, että inositoli 5-pyrofosfaatti (PP-IP5) voi estää DNA korjaantumisen ja siten edistää apoptoosia syövän hoidossa herkistämällä syöpäsoluja.
It is possible that InsP5PP, which contains a pyrophosphate linkage, may function as an ... InsP5PP may inhibit DNA repair and thereby promote apoptosis.
INOSITOLI hexacisfosfaattikinaasi 2 (IP62K) säätelee stimuloivasti IFN-beeta ja gammasäteilyn apoptoottista vaikutusta kaspaasi-8 tietä.
LÄHDE : Bei H Morrison,1 Joseph A Bauer et al. Inositol hexakisphosphate kinase 2 sensitizes ovarian carcinoma cells to multiple cancer therapeutics
Oncogene. 2002 March 14; 21(12): 1882–1889. Oncogene. Author manuscript; available in PMC 2007 October 29. USA.
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2043497
Tutkijat ovat identifioineet inositolihexacisfosfaattikinaasi-2:n ( IP6K2) mahdollisena apoptoosin säätelijänä.
Jos tätä entsyymiä IP6K2 ilmenee ylimäärin, lisääntyy apoptoosi, jota indusoi interferoni beeta (IFN-beeta) ja sytotoksiset agenssit ( tutkittuna ovariaalikarsinoomasoluissa).
IP6K2 tekee inositolipolyfosfaateista IP6 ja IP5 vastaavia pyrofosfaatteja.
IP6K2_HUMAN, Q9UHH9
Tämän entsyymin funktio on muuttaa IP6 muotoon ”IP7”, joka kuitenkin pitää ilmaista eri tavalla, koska inositolissa on vain 6 kohtaa, johon lisäliitos voidaan tehdä. Viidessä on yksi ja yhdessä on kaksi fosfaattia: PP-IP5 on tulos, diphosphoinositol pentakisphosphate(InsP7/PP-InsP5).
Mutta sama entsyymi voi muuttaa myös IP5 molekyylin muotoon PP-IP4, lisäämällä fosfaatin 2- asemaan inositolirengasta. (1,3,4,5,6-pentakisphosphate (InsP5) muotoon PP-InsP4)
Entsyymin muita nimiä:
ATP:1D-myo-inositol-hexakisphosphate phosphotransferase
InsP6 kinase 2
2, 3P(i)-uptake stimulator
3Pi uptake stimulator
2, inositol hexakisphosphate kinase 2
1, 2 inositol hexaphosphate kinase 2 1,
GEENI koodaa proteiinia, joka kuuluu IPK-perheeseen ( inositolifosfokinaasien perheesen) ja tekee yllä olevat muutokset IP6 ja tiettyyn IP5 molekyyliin. Se voi vaikuttaa stimuloimalla IFN-beetan kasvua suppressoivia ja apoptoottisia ominaisuuksia( Tämä on havaittu ovariaalisyövästä) Alternatiivinen pilkkoutuminen geenistä johtaa multippeleihin transkripteihin, jotka koodaavat eri isoformeja.
http://www.genecards.org/cgi-bin/carddisp.pl?gene=IP6K2
ESIMERKKI:
Tässä allaolevassa tutkimuksessa vertailtiin sekä IFN-beeta vaikutusta että gamma- säteilytystä ovariaalisyöpäsolukuoleman (apoptoosin) indusoijina ja tutkijat osoittivat että IP6K2 expressio herkisti tuumorisolut näille kummallekin vaikutukselle.
Jos säteilyttämättömiin syöpäsoluihin transfektoitiin IP6K2, muodostui vähemmän syöpäkolonioita kuin säteilyttämättömiin pelkkää vektoria ilmentäviin syöpäsoluihin
IP6K2 yliexpressoituminen aiheutti lisääntyvää radiosensitiivisyyttä ja se ilmeni alentuneena syöpäkolonioitten muodostumisena. ( CFU= colony forming units)
Sekä IFN-beeta että säteilyttäminen indusoivat kaspaasi-8 tekijää.
http://www.researchapoptosis.com/apoptosis/images/chart-ie.gif
{kind=link}
IFN-beeta ( mutta ei gammasäteily), indusoi syöpäsoluissa TRAIL.
Gammasäteily, mutta ei IFN beeta, indusoi DR4mRNA.
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah&part=A8980&rendertype=figure&id=A8987
Ne apoptoottiset vaikutukset , mitkä tulivat IFN-beetasta ja gammasäteilystä, blokeerautuivat kun expressoitui dominantti negatiivinen mutantti DR5A reseptoria ( death receptoria 5) tai Bcl-2:ta. http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah&part=A8980&rendertype=figure&id=A8987
Kaspaasi -8 mRNA:n indusoituminen oli paljon selvempää niissä soluissa joissa ilmeni IP6K2 verrattuna pelkkää vektoria ilmentäviin soluihin.
http://www.bioscience.org/2007/v12/af/2281/fig1.jp
{kind=link}
DISCUSSION:
In ovarian carcinoma cell lines, IFN-β and γ-irradiation induce apoptosis using similar but not identical pathways. In sensitive cells, IFN-β upregulates TRAIL, and to a lesser extent, caspase 8 (Figure 5a). IFN-β-induced apoptosis has been associated with TRAIL induction in several different tumor cell lines (Chawla-Sarkar et al., 2001; Zhang et al., 1999). γ-irradiation clearly upregulates caspase 8 and DR4 mRNA (Figure 5b) as well as caspase 8 activity (Figure 6a). This is consistent with our previous study showing induction of apoptosis following ionizing radiation (Gong and Almasan, 2000). Transcriptional induction of DR4 following radiation has also been reported in lung carcinoma cell lines (Guan et al., 2001). Our studies are consistent with others, who have described caspase 8 activation following radiation of Jurkat cells (Belka et al., 2000). In three different ovarian carcinoma cell lines, we observed no induction of TRAIL following radiation treatment. This is in contrast to hematologic cells in which both TRAIL and Fas upregulation has been described (Gong and Almasan, 2000). Combination therapy utilizing TRAIL and cytotoxic agents (cisplatin, doxorubicin, or paclitaxel) has been proposed, in an effort to overcome the chemoresistance that usually develops following treatment of women with ovarian carcinoma (Cuello et al., 2001). The p53 status of ovarian carcinoma cells did not correlate with sensitivity to death inducers. In fact, cell lines with mutant p53 (NIH-OVCAR-3, Caov-3) were more sensitive to IFN-β and γ-irradiation than cells containing wild-type p53 (Hey).
We found that DR5Δ protects against apoptosis initiated by both IFN-β and γ-irradiation, confirming that TRAIL : DR-mediated signaling occurs following treatment with both death inducers. Thus, downstream effects of TRAIL and one of its receptors can be blocked by DR5Δ. Combination treatment with both modalities resulted in additive, rather than synergistic antiproliferative effects (Figure 4b). Such additivity is consistent with upregulation of the ligand TRAIL and its receptor DR4, components of the same apoptotic pathway. DR4 and DR5, both agonistic receptors for TRAIL, form homomeric and heteromeric complexes upon ligand binding (Kischkel et al., 2000). Recent reports indicate that ligation of DR4 (Sprick et al., 2000) and DR5 (Bodmer et al., 2000) lead to caspase 8 activation, using a signaling paradigm that is similar, but not identical, to those induced by TNF-α and FasL. Caspase 8 is activated as part of the death-inducing signaling complex (DISC). We show that caspase 8 activity was induced more strongly by radiation compared to IFN-β (Figure 6a,b). Yet caspase 8 activity following combination treatment with IFN-β and γ-irradiation was no greater than that following radiation alone (not shown). It is possible that in NIH-OVCAR-3 cells, caspase 8 activity is already maximized following radiation.
Overexpression of IP6K2 enhanced the degree of apoptosis induced by several different stimuli, including chemotherapeutic drugs, IFNs, and γ-irradiation. Expression of the dominant negative SUB mutant interferes with kinase activity (Morrison et al., 2001) and abrogates death induced by all these agents. IP6K2 protein levels are elevated following IFN-β and irradiation (Figure 1). These findings suggest that IP6K2 functions as a positive regulator of cell death pathways. The mechanism by which this inositol phosphate kinase promotes apoptosis is unclear. IP6K2 converts IP6 to InsP5PP. It is possible that InsP5PP, which contains a pyrophosphate linkage, may function as an energy source that facilitates DISC assembly or signal transduction to the caspase cascade. Proteins that bind IP6 include the clathrin assembly proteins AP-2, AP-3, and AP-180 (involved in membrane fusion) (Norris et al., 1995; Voglmaier et al., 1992; Ye et al., 1995), the synaptotagmins (involved in exocytosis of synaptic vesicles) (Mehrotra et al., 2000) and arrestin (which acts as an adapter during endocytosis) (Gaidarov et al., 1999). All these proteins are closely associated with the plasma membrane. Studies in yeast have suggested that the higher inositol phosphates play a role in mRNA export from the nucleus (Saiardi et al., 2000; York et al., 1999). Finally, IP6 was found to specifically stimulate repair of double-strand breaks in DNA by the process of nonhomologous end-joining in vitro (Hanakahi et al., 2000). If the functions of InsP5PP and IP6 prove to be antagonistic, InsP5PP may inhibit DNA repair and thereby promote apoptosis.
DNA korjausproteiinin MGMT estäminen syöpähoidossa
MGMT on DNA:n korjausproteiini ja kuuluu Direct Reversal –korjausjärjestelmään.
(Engl. O6- alkylguanine DNA alkyl transferase (AGT) repair
AGT repair is a DNA repair protein encoded by MGMT (O6-methylguanine DNA methyltransferase) removes DNA adducts from O6 methylguanine. The failure in the repair system results in G to A transition and DNA strand break observed in hematopoietic tumors, breast, ovarian and retinoblastomas ). www.molecular-cancer.com/content/6/1/26
Jos MGMT ei toimi ei pystytä DNA:n sisällä korjaamaan alkyloitunutta G -emästä ja voi tapahtuu luennassa G-A transition(eikä normaali G -C linjan luenta) ja DNA:n säikeen katko.
Mutta syöpäkasvaimen suhteen taas koetetaan saada aikaan tällaista DNA-vauriota syövässä, jotta se saataisiin pysähtymään solu solulta niiden alkaessa jarruttaa solusykliä.
Suomennosta abstraktista.
Eräs vaikea syöpälaji haimasyöpä on hyvin kestokykyinen sellaisia syöpälääkkeitä vastaan, jotka alkyloivat DNA:ta. Yksi syy on, että haimasyövässä on korkeat pitoisuudet tätä MGMT -entsyymiä, DNA:n korjausproteiinia.
(MGMT= DNA repair protein, O6-methylguanine-DNA methyltransferase).
Miten voidaan saada syöpäkudos herkemmäksi (alkyloivalle) sytostaattiselle lääkeaineelle?
Voidaan koettaa inaktivoida tämä MGMT-entsyymi jollain inhibiittorilla, kuten tutkijat abstraktissaan kertovat.
He käyttivät erästä MGMT inhibiittoria BG (O6-benzyyli-guaniinia).
Alkyloivat aineet , jolla syöpäsoluja tutkittiin, olivat tässä carmustine (BCNU) ja temozolomide (TMZ).
Mikä oli p53 osuus?
Tutkijat huomasivat, että jos p53 tekijä( genomin suojelija) oli mutatoitunut, kombinaatioterapia TMZ +BG tehosi huonommin kuin silloin, jos p53 oli kunnossa.
TMZ + BG aiheutti solusyklin pysähtymän tekemällä DNA-vaurion ja solut joutuivat apoptoosiin, varsinkin haimasyöpäkudoksessa. Apoptoosi välittyi p53 tietä, sillä oletetaan , että p53 kontrolloi solusyklin jarruttumista (arrest) joko G1/S tai G2/M kontrollipisteissä antamalla pro-apoptoottisen vasteen.
KTS kuva: P53 kohdeproteiinit, joiden kautta p53 vaikuttaa apoptoosin:
http://www.nature.com/nrc/journal/v1/n1/images/nrc1001-068a-f1.gif)’
{kind=link}
Tutkijat selvittivät edelleen mekanismia, jolla funktionaalinen p53 tekijä vaikutti omalta osaltaan korjaamattomien O6-metyyliguaniinien toksisuuteen.
Mutatoitunutta p53 tekijää Mut-P3 sisältänyt solulinja (Capan-1) sisältää korkeat basaalipitoisuudet p53, p21, Bax ja Bcl-2 ja näitten proteiinien pitoisuudet pysyvät muuttumattomina, kun käsitellään kombihoidolla BG + TMZ.
Mutta jos käsiteltiin sitä linjaa jossa on normaali p53 (Capan-2 linjaa) osoittautuu tapahtuvan merkitsevä indusoituminen genominsuojelijatekijässä p53 ( joka on tuumorisuppressori) sekä p21 tekijässä samaisella kombikäsitelyllä.
Eräällä TUNLl-menetelmällä voitiin osoittaa kombikäsittelyn (TMZ+BG) jälkeen, että normaalilla p53 tekijällä saatiin aikaan huomattavasti suurempi, kaksinkertainen apoptoottinen indeksi, verrattuna mutantti-p53-linjaan. Tämä vaste korreloi BAX-expression indusoitumiseen.
Merkitsevän tärkeää tässä on p53 proteiinin vaikutus mismatch-korjaus (MMR) proteiinin hPMS2 esiin indusoituminen, joka taas osallistuu jälkipäätöksiin tunnistettuaan O6-metyyliguaniini mismatch-emäsparin esiintymisen ja voi johtaa solun kuolemaan.
(KTS MMR http://www.nature.com/cr/journal/v18/n1/full/cr2007115a.html )
(MMR komponentit: :
http://www.nature.com/cr/journal/v18/n1/fig_tab/cr2007115t1.html#figure-title
Basaaliset pitoisuudet hPMS2, p21 ja BAX-proteiineja oli merkitsevästi matalammat kun p53 oli normaali verrattuna mutantti Mut-p53-soluihin
Kombiterapian jälkeen(BG+TMZ) säätyi ylös hPMS2 normaalin p53 omaavissa soluissa, kun taas mutantti-53 soluissa sen pitoisuudet laskivat (=MMRjärjestelmä osallistuu jatkossa)
Ylläolevan kanssa yhdenmukaisesti: solusyklin pysähtymän indusoiminen PI3K/CDK inhibiittorilla (7-OH-staurosporin) mutantti-p53:n omaavat haimasyöpäsolusiirrännäiset johtivat koe-eläimessä selvästi tuumorikasvun viiveeseen vasteena kerta käsittelylle TMZ+BG.
(Solusyklin pysähtymä sinänsä voi olla p53:sta riippumatonkin:
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah&part=A20842)
LÄHDE: Sabapathi Sathishkumar, Seema Gupta, Mansoor M. Ahmed and Demetrius M. Kokkinakis Role of p53 and cell cycle arrest in the sensitization of pancreatic tumors to temozolomide University of Kentucky, Lexington, KY and University of Pittsburgh, Pittsburgh, PA
Ref: Kokkinakis DM, Ahmed MM, et al. Sensitization of pancreatic tumor xenografts to carmustine and temozolomide by inactivation of their O6-Methylguanine-DNA methyltransferase with O6 benzylguanine or O6-benzyl-2’-deoxyguanosine. Clin Cancer Res. 2003 Sep 1;9(10 Pt1):3801-7.
http://aacrmeetingabstracts.org/cgi/content/abstract/2004/1/1299DNA:n korjaantuminen suoraa entsyymillä, uutisia
Uutiset näistä entsymaattisesta DNA korjauksesta , Direct reversal
Suora entsymaattinen DNA:n korjaus
DNA voi korjaantua siis aivan suoraan entsymaattisesti ilman monimutkaisemman replikaatiofunktion osuutta.
Näitä on 4 erilaista mekanismia ( Kts. edellinen artikkeli) . PubMed antaa 3 artikkelia, jossa kuvataan näistä uusia asioita Hakusana DNA Repair Direct Reversal
LÄHDE 1. Bakteeri-itiöiden sitkeyden taustaa
FOTOLYAASI
Chandra T, Silver SC et al. Spore photoproduct lyase catalyzes specific repair of the 5R but not the 5S spore photoproduct. J Am Chem Soc. 2009 Feb 25;131(7):2420-1.Related Articles
Bakteerien itiöt ovat aivan ihmetyttävän kestäviä kaikenlaista kemiallista ja fysikaalista stressiä kohtaan, myös UV-sädetystä vastaan. Epätavallinen UV-säteilyn kestävyys bakteerien itiöissä johtuu niiden ainutlaatuisesta itiö-DNA:n fotokemiasta, joka aiheuttaa, että kertyy 5-thyminyyli-5,6-dihydrothymiiniä (SP-materiaa, itiöitten valotuotetta, spore photoproduct) ja sitä liittyneenä akkumuloituneen vaurion tehokkaasti korjaavaan SPL-entsyymiin, fototuotteen lyaasiin, fotolyaasiin.
SPL on jäsen radikaaleissa AdoMet superperheen entsyymeissä ja se hyödyntää rauta-rikki-kertymiä ja S-adenosyylimetioniinia (SAM) korjatakseen SP tuotteen mainitulla ”suoralla reversiolla” ( direct reversal mechanism) kiskomalla H-atomin thymiinidimeerin C6 hiilestä.
Bakteerien itiöitä UV-säteilytettäessä voi muodostaa periaatteeessa kaksi SP-diastereomeeriä (5R ja 5S) , mutta vain 5R-konfiguraatiota on mahdollista muokata lähellä olevista thymiineistä DNA:n kaksoishelixistä riippuen DNA rakenteen asettamista rajoituksista . Mutta 5S konfiguraation muodostuminen on mahdollinen vähemmin täsmällisissä DNA-rakenteissa tai säikeitten välisissä poikkisidoksissa.
Tutkijat analysoivat SP stereokemialliset erot ja osoittivat, että fotolyaasientsyymi SPL korjaa SP-tuotteessa olevat 5R isomeerit ainoastaan. SPL –entsyymin substraattina on siis 5R-SP eikä 5S-SP ja tämä tieto pitää yhtä sen havainnon kanssa, että kun UV-säteilytetään bakteerien itiöiden DNA:ta in vivo, tuottuu vain 5R-SP isomeeriä.
LÄHDE: 2. Tavallisimmat alkylaatiot mA ja mC
Leiros I, Nabong MP et al. Structural basis for enzymatic excision of N1-methyladenine and N3-methylcytosine from DNA. EMBO J. 2007 Apr 18;26(8):2206-17. Epub 2007 Mar 29.
Yksinkertaisiin DNA-säikeisiin toksiinien ja mutageenien aiheuttamsta alkylaatioista on tavallisimmat tyypit
N(1)-metyladeniini(m(1)A)
N(3)-metyylicytosiini (m(3)C)
Sekä bakteereilla että ihmisillä on osoitettu m(1)A ja m(3)C alkylaatioitten korjautuvan AlkB-välitteisellä oksidatiivisella demetylaatiolla, mikä on suora DNA-vaurion palauttava mekanismi (direct reversal).
Mutta Archea-lajeissa ei ole todettu AlkB-geeni-homologeja. Miten niillä DNA pystyy korjautumaan?
Tutkijat osoittivat, että lajilla Archaeoglobus fulgidus, nuo mainitut alkylaatiot m(1)A ja m(3)C korjaantuivat AfAlkA base excision repair glycosylase-menetelmällä. Siis tuossa kolmannessa elämän domaanissa toimisi toinen mekanismi. Lisäksi havaittiin, että AfAlkA aiheutti runsaan 1,N(6)-ethenoadenine excision .
Tutkijat esittivät AfAlkA kiderakenteen luonnehtien sitä myös pistemutaatioilla ja esittivät täten uuden löydön base excision- aktiivisuudesta
LÄHDE 3. DNA:n korjautumisessakin on kunto tärkeä
Kreklau EL, Limp-Foster M, Liu N, Xu Y, Kelley MR, Erickson LC.A novel fluorometric oligonucleotide assay to measure O( 6)-methylguanine DNA methyltransferase, methylpurine DNA glycosylase, 8-oxoguanine DNA glycosylase and abasic endonuclease activities: DNA repair status in human breast carcinoma cells overexpressing methylpurine DNA glycosylase. Nucleic Acids Res. 2001 Jun 15;29(12):2558-66.
DNA:n korjaamiskyky (DNA repair status) on tärkeä seikka mutageneesissa, carsinogeneesissa ja genotoksisten aineitten kestokyvyssä. Koska DNA:n korjaamisprosessit käsittävät monia entsymaattisia askeleita, on solun DNA:n reparatiivisen kyvyn ymmärtämiseksi tarvinnut kehittää useita tutkimusmenetelmiä.
Tässä tutkijat esittävät uuden kehittelemänsä koeputkimenetelmän, jolla voidaan määrittää yksinkertaisista solu-uutteista kvantitatiivisesti alkylaatioitten korjaantuminen MGMT tietä ja BER-tietä , jossa on mukana MPG glykosylaasientsyymi, hOGG1 glykosylaasientsyymi ja hiivan ja ihmisen abaasinen endonukleaasi ( APN1 ja vastaavasti APE/ref/1
MGMT= O(6)-methylguanine DNA methyltransferase , korjaa poistamalla metyyliä
BER= base excision repair, korjaa poistamalla virheellisen emäksen
MPG= methylpurine DNA glycosylase
hOGG1= human 8-oxoguanine DNA glycosylase (hOGG1)
APN1= yeast abasic endonuclease
APE/ref-1 = human abasic endonuclease
Tämä lähestymistapa käsittää sellaisen solu-uutteen valmistuksen, jossa on yleispuskurissa kaikkia DNA-korjausprosessiin osallistuvia proteiineja aktiivina ja käytetään fluorometrisesti merkattuja stabiileja oligonukleotidisubstraatteja, joissa on jokaiselle DNA-korjausproteiinille spesifisiä DNA-leesioita.
Tämä metodi tekee mahdolliseksi määrittää metylaatio ja BER-kapasiteetit nopeasti pienestä määrästä alkunäytettä. Lisäksi fluorometristen oligonukleotidien stabiliteetilla vältetään substraatin muuntuvuus, mitä jatkuva radioaktiivinen merkkaus aiheuttaa
(Kts. mahdolliset metylaatiokohdat
http://www.mgmt-agt.net/Methylation%20sites2.jpg)
{kind=link}
Tässä raportissa tekniikkaa sovellettiin ihmisen rintasyöpäsoluihin (MDA-MB231), jotka ilmentävät ylimäärin ihmisen MPG entsyymiä, jotta voitiin mitata, muuttaako BER- järjestelmän alkuvaiheen ylössäätyminen aktiivisuutta muissa valituissa BER (hOGG1) ja APE (ref-1 korjausjärjestelmissä tai suoraankorjaavaa (direct reversal) ( MGMT) aktiivisuutta.
Kommenttini:
Olen jo pitempään arvellut, että proteiinien ja antioksidanttien saantiin tulisi kiinnittää huomio DNA repair- statuksen ( tasapainon) kannalta, EIKÄ vain siltä kannalta, mitä keho erittää jännöstyppeä ja mikä on minimi tarve, että ei jouduta negatiiviseen typpitasapainoon jostain tuntemattomasta essentiellistä seikasta.
DNA:n korjaantuminen suoraa entsyymillä
Taustaa tekijöistä jotka vahingoittavat DNA:ta
Röntgensäteet, vapaat radikaalit, alkyloivat aineet ja spontaanireaktiot
aiheuttavat urasiilin muodostusta, abaasisen kohdan, 8-oxoguaniinia, ja yhden säikeen katkeamaa ( single strand break).
Näitä korjaa BER –järjestelmä , joka tarvitsee DNA-glykosylaaseja, APE1 endonukleaasia, DNApolymeraasi betaa ja XRCC1-DNA-ligaas1:tä. BER järjestelmässä on varsinaista DNA:n synteesiä jo mukana, mutta on olemassa myös suoran reversion järjestelmä, missä ei ole varsinaista DNA-synteesiä (replikaatiota) vielä mukana, vaan asia on entsymaattisella tasolla nikkaroimista.
”Direct Reversal” on tämän suoran korjauksen nimi
Tunnetaan neljä eri DNA-vaurion suorakorjausjärjestelmää.
- Photolyase, fotolyaasientsyymi korjaa UV-säteilyn aiheuttaman vaurion pyrimidiinidimeerissä valosta riippuvassa reaktiossa. Tämä korjaus tapahtuu bakteerilla, mutta ei ihmisellä.
- DNA ligaasi korjaa yksinkertaisen DNA mutkan.
Ihmissoluissa on useita DNA-ligaaseja, joilla on erilaisia tehtäviä niin DNA::n korjauksessa kuin monimutakisemmassa replikaatiossa. (Esim. Polymeraasi beeta on interaktiossa DNA-ligaasi I:n kanssa suoraan ja epäsuoralla tavalla taas XRCC1:n välityksellä . (1996). DNA-ligaasi III alfa muodostaa stabiilin kompleksin XRCC1:n kanssa ja on interaktiossa myös PARP1 ja PARP2:n kanssa.
Nämä molemmat proteiinit ovat interaktiossa keskenään ja polymeraasi beetan kanssa ja samalla tekevät linkin DNA ligaasi IIIalfan ja Base Excision Repair (BER)- ja single -strand break repair- mekanismeihin, koska XRCC1 ja PARP ovat jälkimmäisessä osallisia.
Suurin osa short patch repair- tapahtumista saadaan täydellisesti suoritetuksi DNA-ligaasi IIIalfa/XRCC1- kompleksilla.
DNA-ligaasi IIIalfa ei ole assosioitunut replikaatiofokuksiin eikä ole BER:ssä mukana , jos se tapahtuu lähellä replikaatiohaarukkaa) .
Poikkeavat metylaatiot DNA:ssa korjataan BER-systeemillä ja suoralla korjauksella joko metyylitransferaasilla tai oksidatiivisella demetylaasilla, joka on AlkB-tyyppiä.
- Metyylin poistajat, MGMT, O-metylguanine-DNA-methyltransferase
http://www.mgmt-agt.net/mgmt-function.JPG
{kind=link}
Tämä entsyymi siirtää metyylin tai muun yksinkertaisen alkyyliryhmän O6-alkyyliguaniinista (O6-mG) tai O4-alkyylithymiinistä omaan cysteiiniinsä, jolloin entsyymi itse inaktivoituu vapauttaessaan nukleotidin. Tämä entsyymi luo soluissa resistenssiä Guaniinia O6-alkyloivien aineitten sytotoksisuutta vastaan. (Toisaalta solut voidaan tehdä herkiksi alkylaatiolle alentamalla tämän MGMT-entsyymin vaikutusta).
Jos O6-mG ei korjaannu, seuraa ”mismatch” thymiiniin päin (T) ja silloin joutuu mismatch repair ( MMR)- systeemi triggeröitymään ja apoptoosi voi seurata.
- Alkyylin irrottajat AlkB, hABH12, hABH3 ( human AlkB homologue)
Alkyloitunut DNA voi korjautua siten, että alkyyli voidaan saada irrotettua. Alfa-KG:sta ja raudasta riippuva oxygenaasi hydroxyloi DNA-emäksen alkyyliryhmän, jolloin alfa-KG muuttuu meripihkahapoksi ja alfa-KG:n C5 irtoaa hiilidioksidina Hydroksyloitu metyyli vapautuu spontaanisti formaldehydinä
Tällä korjauksella irtoaisi CH3- ryhmä 1-meA.sta tai 3-meC.stä korjaten adeniinin ja cytosiinin DNA.ssa ja RNA.ssa. Tekijää tai entsyymiä sanotaan AlkB (DNA and RNA), tai hABH12 (DNA) tai hABH3 (DNA and RNA).
Tästä on uutta tietoa vuodelta 2008:
Yang et al. [Nature 452 (2008)
AlkBrakenne DNA-kompleksissa on voitu määrittää, samoin humaani AlkB homologi 2 rakenne (hABH2) käyttämällä crosslinking-strategiaa, jolla on DNA-proteiinikompleksi stabiloitu. AlkB-proteiineilla on samanlainen katalyyttinen domaani, mutta DNA-tunnistusmotiivit ovat erilaisia. AlkB tekee kontaktin pääasiassa vaurioituneesen säikeeseen, mutta hABH2 tekee useita kontakteja molempien säikeitten kanssa . hABH2 luiskauttaa ulos vaurioituneen emäksen ja täyttää tyhjän paikan hydrofobisella aminohappotähteellä, samaan tapaan kuin DNA-glykosylaasit , essentiellisti niin, että DNA-kaksoishelixrakenne ei kierry. Sitävastoin AlkB puristaa niitä emäksiä yhteen, mitkä sijaitsevat irrotettavan emäksen kyljissä siten että irtiluiskahtava emäs joutuu pinon päälle. Tämä odottamaton irrotuskeino ja ero AlkB ja hABH2:n välillä DNA-säiekontaktissa selittää, miksi niillä on eri prioriteetit single strand break ja DSB DNA vaurioissa vastaavasti.
http://www.ncbi.nlm.nih.gov/bookshelf/picrender.fcgi?book=eurekah&part=A79604&blobname=ch4177f2.jpg
{kind=link}
http://www.ncbi.nlm.nih.gov/bookshelf/picrender.fcgi?book=eurekah&part=A79604&blobname=ch4177f4.jpg
{kind=link}
onsdag 3 juni 2009
Kuinka pitkä on kehon solujen kaikki DNA?
DNA:n rakenneosaset: A,T,G,C
DNA:ssa esiintyvät emäkset ovat : Adenine (A), thymine (T), cytosine (C) ja guanine (G)
(GG = Global Genome)
Ihmisen Genomi on diploidinen ja sisältää 22 kromosomiparia ja kaksi sukupuolikromosomia, XX naisilla ja XY miehillä.
Solun DNA-pitoisuus:
DNA pitoisuus genomissa on 3 x 10*9 bps / haploidi genomi
3 000 000 000 emäsparia
Bp on base pair, emäspari
Jokaisen emäsparin välillä on 3,4 Å mitta.
Joten yhden solun DNA:ta on 2 metriä / diploidi genomi.
Kun ihmisellä on kehossaan soluja noin 10*exp. 13, siitä seuraa että nukleotideja on ihmisessä 3 x 10 E22 eli DNA mittaa tulee 2 x 10E10 km.(20 000 000 000 km).
Auringon ja maan etäisyys on 147 100 000 km.
DNA-yhteispituus ihmiskehon soluissa on siis noin 130 kertaa etäisyys auringosta maahan.
DNA:n itsensätoistamisessa virheen mahdollisuuksien määrä on tähtitieteellinen. Joten se elämisen järki, joka pakkautuu koodiin ja integroituna toteutuu siten, että tulos on elävä, toimiva, ajatteleva, järkevä, luova ja lisääntyvä ihminen, on mielestäni ihme.
Kehon ainoa keino välttyä liialta perinnöllisesltä mutatoitumiselta, lienee erikoistaa tärkeät solut ja pysäyttää ne G0-tilaan, mikä on parempi kuin mutaatioriski, ja kompromissinä tälle suojatekijälle on rajoittunut ihmiskunnan ikä.
On olemassa järjestelmä, joka pitää solua G0(zero) tilassa, joten koko genomi ei toista joka hetki tuota koko matkaa, sitä 130 maan ja auringon väliä, vaan välttämättömän, essentiellin osan siitä ja senkin synkronisesti eri kohdissa S- faasia. Jos genomi toistaisi koko itsensä jatkuvasti, ihmisen energiantarve olisi aivan suunnaton, itse asiassa täysi mahdottomuus toteuttaa.
Kun ajattelee DNA:n luonnetta, että se lisääntyy uskollisesti itseään kopioimalla, tämä asettaa soluille erittäin suuret vaatimukset, jotta noin pitkä geneettinen informaatio voi vuodesta toiseen toistua virheettömästi. DNA:n uusiutumisen virheistä ja hyvistä sattumista summautuu luonnollisesti ihmiskunnan iän rajat ja DNA:n itsensäkorjaamiskyvyn heikentymisestä syntyy sairaustiloja kuten syöpiä. Ihmiskunnalle on eduksi hankkia lapsia kohtalaisien nuorella iällä. pojasta polvi paranee, sanotaan. Elämänvoimat valitsevat parempaa ja elinkykyisempää esiin.
Jotta noin mittavan DNA:n uusiutumiskopioinnit onnistuisivat onnellisesti, täytyy kehon joka päivä olla ravitsemukseltaan, hapensaanniltaan ja miljöötekijöiltään otollisessa tilassa. (DNA:n kannalta katsoen se ehkä voisi tuottaa 180 vuoden elinikää, mutta elinkunto rajoittaa elinikää noin 70-80 maille. Monet uudet sairaudet yhteiskunnassa vikuttavat myös globaalin genomin stabiliteettia ja täten elinikää.
Nobelin palkinnot saaneet keskustelivat takavuosina Tukholman Bernadotte-salissa eliniästä, ja eräs ekonomian tiedemies totesi, että ekonomisinta olisi 300 vuoden elinikä, kun kerran nuoret koulutuksen hankkivat pääsevät tuottoisaan elämään vasta noin 30 vuotiaina ja sitten voivat toimia vain. seuraavat 30 v. nykyisellään.
DNA:ssa esiintyvät emäkset ovat : Adenine (A), thymine (T), cytosine (C) ja guanine (G)
(GG = Global Genome)
Ihmisen Genomi on diploidinen ja sisältää 22 kromosomiparia ja kaksi sukupuolikromosomia, XX naisilla ja XY miehillä.
Solun DNA-pitoisuus:
DNA pitoisuus genomissa on 3 x 10*9 bps / haploidi genomi
3 000 000 000 emäsparia
Bp on base pair, emäspari
Jokaisen emäsparin välillä on 3,4 Å mitta.
Joten yhden solun DNA:ta on 2 metriä / diploidi genomi.
Kun ihmisellä on kehossaan soluja noin 10*exp. 13, siitä seuraa että nukleotideja on ihmisessä 3 x 10 E22 eli DNA mittaa tulee 2 x 10E10 km.(20 000 000 000 km).
Auringon ja maan etäisyys on 147 100 000 km.
DNA-yhteispituus ihmiskehon soluissa on siis noin 130 kertaa etäisyys auringosta maahan.
DNA:n itsensätoistamisessa virheen mahdollisuuksien määrä on tähtitieteellinen. Joten se elämisen järki, joka pakkautuu koodiin ja integroituna toteutuu siten, että tulos on elävä, toimiva, ajatteleva, järkevä, luova ja lisääntyvä ihminen, on mielestäni ihme.
Kehon ainoa keino välttyä liialta perinnöllisesltä mutatoitumiselta, lienee erikoistaa tärkeät solut ja pysäyttää ne G0-tilaan, mikä on parempi kuin mutaatioriski, ja kompromissinä tälle suojatekijälle on rajoittunut ihmiskunnan ikä.
On olemassa järjestelmä, joka pitää solua G0(zero) tilassa, joten koko genomi ei toista joka hetki tuota koko matkaa, sitä 130 maan ja auringon väliä, vaan välttämättömän, essentiellin osan siitä ja senkin synkronisesti eri kohdissa S- faasia. Jos genomi toistaisi koko itsensä jatkuvasti, ihmisen energiantarve olisi aivan suunnaton, itse asiassa täysi mahdottomuus toteuttaa.
Kun ajattelee DNA:n luonnetta, että se lisääntyy uskollisesti itseään kopioimalla, tämä asettaa soluille erittäin suuret vaatimukset, jotta noin pitkä geneettinen informaatio voi vuodesta toiseen toistua virheettömästi. DNA:n uusiutumisen virheistä ja hyvistä sattumista summautuu luonnollisesti ihmiskunnan iän rajat ja DNA:n itsensäkorjaamiskyvyn heikentymisestä syntyy sairaustiloja kuten syöpiä. Ihmiskunnalle on eduksi hankkia lapsia kohtalaisien nuorella iällä. pojasta polvi paranee, sanotaan. Elämänvoimat valitsevat parempaa ja elinkykyisempää esiin.
Jotta noin mittavan DNA:n uusiutumiskopioinnit onnistuisivat onnellisesti, täytyy kehon joka päivä olla ravitsemukseltaan, hapensaanniltaan ja miljöötekijöiltään otollisessa tilassa. (DNA:n kannalta katsoen se ehkä voisi tuottaa 180 vuoden elinikää, mutta elinkunto rajoittaa elinikää noin 70-80 maille. Monet uudet sairaudet yhteiskunnassa vikuttavat myös globaalin genomin stabiliteettia ja täten elinikää.
Nobelin palkinnot saaneet keskustelivat takavuosina Tukholman Bernadotte-salissa eliniästä, ja eräs ekonomian tiedemies totesi, että ekonomisinta olisi 300 vuoden elinikä, kun kerran nuoret koulutuksen hankkivat pääsevät tuottoisaan elämään vasta noin 30 vuotiaina ja sitten voivat toimia vain. seuraavat 30 v. nykyisellään.
Kaikki DNA:n korjaantumisjärjestelmät
Tämä kuva on nettilähteestä
http://herkules.oulu.fi/isbn9514267141/html/graphic11.png
Fytiiniaineenvaihdunnan osuus on pohdittava erikseen jokaisessa prosessissa.
Tällä hetkellä olen katsomassa NHEJ ja HR kaksoisvaurion korjaantumista.
3.6.2009 9:56
http://herkules.oulu.fi/isbn9514267141/html/graphic11.png
{kind=link}
Fytiiniaineenvaihdunnan osuus on pohdittava erikseen jokaisessa prosessissa.
Tällä hetkellä olen katsomassa NHEJ ja HR kaksoisvaurion korjaantumista.
3.6.2009 9:56
tisdag 2 juni 2009
DNA:n korjausjärjestelmä NHEJ
NHEJ
Non-homologous DNA End Joining. Kuvaan yksityiskohtaisemmin tätä järjestelmää, jossa toimii Ku proteiini ja DNA-PK entsyymi.
LÄHDE:
E. Pastwa and J. Blasiak,
Repair of DNA DSBs by NHEJ in mammalian cells.
Reprinted form Acta Biochimica Polonica 50(4), Non-homologous DNA End Joining, 891-908, 2003,
Vuosi 2004 on nettiin tullut tällainen versio, joka selvittää NHEJ- DNA:n kaksoisvaurion (DSB) korjaantumista
http://www.ebi.ac.uk/interpro/potm/2004_7/Page2.htm
MITEN NHEJ TOIMII?
Tämä non-homologinen säikeitten päitten liittyminen toimii kaikenlaisissa soluissa, aivan bakteereistä ihmiseen ja se osallistuu monenlaisiin proseseihin, kuten DNA:n korjaukseen, telomeerien ylläpitoon ja HIV-1 proviruksen kiinnittämiseen ihmisen genomiin DNA:han ja toistosegmenttien Tandem Repeat kiinnittämiseen.
DNA:n DSB Double Strand Break kaksoiskatkon säikeiden liittämiseen osallistuu proteiinisarja, jotka tekevät yhteistyötä, jossa ne saavat suoritettua katkenneiden DNA-päitten synapsoitumisen, trimmauksen ja ligaasilla yhdistämisen.
Eukaryoottisissa soluissa pääasiallisimmat proteiinit, jotka osallistuvat NHEJ-tiehen ovat
DNA-ligaasi IV,
XRCC4,
DNA-PKcs ( DNA:sta riippuvan proteiinikinaasin katalyyttinen alayksikkö),
Ku-proteiini
ja mahdollisesti Artemis.
Jos missä tahansa näistä proteiineista esiintyy puutteita, seikka johtaa yliherkkyyten DNA-DSB vaurioita indusoivia aineita kohtaan, kuten mm jonisoivaa säteilyä kohtaan.
On identifioitu kolme askelta NHEJ-prosessissa.
sitoutuminen DNA:n päihin ja sillan teko.
terminaalisten päätyjen prosessoinnti
liittäminen ligaasilla.
(DNA-dependent protein kinase = DNA-PK).
Ku muodostaa rinkulan, joka ympäröi kaksoisDNA:ta kietoutuen kaksi täyttä kierrosta DNA-molekyylin ympäri. Kun Ku muodostaa sillan murtuneitten DNA-päätyjen välille , se toimii rakenteellisena kannattajana ja oikaisijana suojaten samalla päätyjä hajoamiselta ja estää samalla sekavat sitoutumiset ehjään DNA:han. Ku suoristaa tehokkaasti DNA:ta ja samalla sallii polymeraasien, nukleaasien ja ligaasien päästä vaikuttamaan katkenneen DNA:n päätyihin edistämään päitten liittämistä yhteen.
Heti kun Ku on paikan päällä, se rekrytoi DNA-PK entsyymin katalyyttisen alayksikön (DNA-PKcs), joka on seriini/treoniini-kinaasi ja kuuluu PI3K-perheeseen, jota pidetään signaalimolekyylinä vastaamassa solun stressiin. (phosphoinositide 3-kinase (=PI3K) family)
DNA-PKcs pystyy fosforyloimaan useita tumaproteiineja in vitro, ja näihin kuuluu DNA ligaasi IV ja XRCC4.
DNA ligaasi IV:n ja /tai XRCC4:n fosforylaatiot voivat vaikuttaa Ku-proteiinin ja muiden proteiinien keskinäisiin interaktioihin.
Vaihtoehtoisesti tällainen ss (single stranded) roikkuva säie voidaan trimmata nukleaasin aktiivisuudella. Tässä tehtävässä saattaa olla osuutta Artemis-proteiinilla, joka kuuluu metallo-beetalaktamaasi-perheeseen.
DNA-PKcs entsyymi fosforyloi ja sitoutuu Artemisproteiiniin. Nyt muodostuvlla kompleksilla Artemis/DNA-PKcs arvellaan olevan nukleaasiaktiivisuutta, joka pystyy pilkkomaan irti 5´ ja 3´ DNA-roikkuvat kohdat
XRCC4 toimii säätelyelementtinä, joka stabilisoi DNA-ligaasiIV-entsyymiä, stimuloi sen aktiivisuutta ja ohjaa ligaasia DNA-vauriokohtaan omalla helix- rakenteisella tunnistuskohdallaan ja DNA:ta sitovalla kyvyllään.
On tehty lukuisia tutkimuksia jotta voitaisiin identifioida muitakin NHEJ-järjestelmän komponentteja.
http://www.ebi.ac.uk/interpro/potm/2004_7/Page2.htm

KOMMENTTI
Tässä huomataan, että retrovirus pystyy käyttämään solun DNA:n omaa korjausjärjestelmää ja asentaa proviruksensa DNA-muotoisena tällä järjestelmällä isäntäkehon genomiin, eikä järjestelmä tai muu järjestelmä tunnista vierasta DNA:ta. Tällä tavalla HIV-1 virus on tositaiseksi pandeminen kautta maailman ja tuottaa genomin instabiliteetin.
Tässä alla olevassa kuvassa retrovirus toimii miltei kuten proteiini Ku joka lähtee korjaamaan virhettä, mutta retrovirus tekee virheen isäntäsolun DNA:han. ja leikkaa sen poikki ja sitten ”korjaa ”asettamalla oman itsensä väliin ja herättää korjausjärjestelmän isäntäsolun DNA.n katkaisemalla. Siis (ainakin) NHEJ järjestelmä on retroviruksen orjuuttama.
http://www.nature.com/cdd/journal/v12/n1s/images/4401573f1.jpg

http://www.nature.com/cdd/journal/v12/n1s/full/4401573a.html#fig1
2.6.2009 17:16.Päivitys 4.2. 2015.
Non-homologous DNA End Joining. Kuvaan yksityiskohtaisemmin tätä järjestelmää, jossa toimii Ku proteiini ja DNA-PK entsyymi.
LÄHDE:
E. Pastwa and J. Blasiak,
Repair of DNA DSBs by NHEJ in mammalian cells.
Reprinted form Acta Biochimica Polonica 50(4), Non-homologous DNA End Joining, 891-908, 2003,
Vuosi 2004 on nettiin tullut tällainen versio, joka selvittää NHEJ- DNA:n kaksoisvaurion (DSB) korjaantumista
http://www.ebi.ac.uk/interpro/potm/2004_7/Page2.htm
MITEN NHEJ TOIMII?
Tämä non-homologinen säikeitten päitten liittyminen toimii kaikenlaisissa soluissa, aivan bakteereistä ihmiseen ja se osallistuu monenlaisiin proseseihin, kuten DNA:n korjaukseen, telomeerien ylläpitoon ja HIV-1 proviruksen kiinnittämiseen ihmisen genomiin DNA:han ja toistosegmenttien Tandem Repeat kiinnittämiseen.
DNA:n DSB Double Strand Break kaksoiskatkon säikeiden liittämiseen osallistuu proteiinisarja, jotka tekevät yhteistyötä, jossa ne saavat suoritettua katkenneiden DNA-päitten synapsoitumisen, trimmauksen ja ligaasilla yhdistämisen.
Eukaryoottisissa soluissa pääasiallisimmat proteiinit, jotka osallistuvat NHEJ-tiehen ovat
DNA-ligaasi IV,
XRCC4,
DNA-PKcs ( DNA:sta riippuvan proteiinikinaasin katalyyttinen alayksikkö),
Ku-proteiini
ja mahdollisesti Artemis.
Jos missä tahansa näistä proteiineista esiintyy puutteita, seikka johtaa yliherkkyyten DNA-DSB vaurioita indusoivia aineita kohtaan, kuten mm jonisoivaa säteilyä kohtaan.
On identifioitu kolme askelta NHEJ-prosessissa.
sitoutuminen DNA:n päihin ja sillan teko.
terminaalisten päätyjen prosessoinnti
liittäminen ligaasilla.
- DNA-päihin sitoutuminen ja sillanteko ( DNA End-binding and Bridging)
(DNA-dependent protein kinase = DNA-PK).
Ku muodostaa rinkulan, joka ympäröi kaksoisDNA:ta kietoutuen kaksi täyttä kierrosta DNA-molekyylin ympäri. Kun Ku muodostaa sillan murtuneitten DNA-päätyjen välille , se toimii rakenteellisena kannattajana ja oikaisijana suojaten samalla päätyjä hajoamiselta ja estää samalla sekavat sitoutumiset ehjään DNA:han. Ku suoristaa tehokkaasti DNA:ta ja samalla sallii polymeraasien, nukleaasien ja ligaasien päästä vaikuttamaan katkenneen DNA:n päätyihin edistämään päitten liittämistä yhteen.
Heti kun Ku on paikan päällä, se rekrytoi DNA-PK entsyymin katalyyttisen alayksikön (DNA-PKcs), joka on seriini/treoniini-kinaasi ja kuuluu PI3K-perheeseen, jota pidetään signaalimolekyylinä vastaamassa solun stressiin. (phosphoinositide 3-kinase (=PI3K) family)
DNA-PKcs pystyy fosforyloimaan useita tumaproteiineja in vitro, ja näihin kuuluu DNA ligaasi IV ja XRCC4.
DNA ligaasi IV:n ja /tai XRCC4:n fosforylaatiot voivat vaikuttaa Ku-proteiinin ja muiden proteiinien keskinäisiin interaktioihin.
- Terminaalisten päätteitten prosessoiminen (Terminal End Processing)
Vaihtoehtoisesti tällainen ss (single stranded) roikkuva säie voidaan trimmata nukleaasin aktiivisuudella. Tässä tehtävässä saattaa olla osuutta Artemis-proteiinilla, joka kuuluu metallo-beetalaktamaasi-perheeseen.
DNA-PKcs entsyymi fosforyloi ja sitoutuu Artemisproteiiniin. Nyt muodostuvlla kompleksilla Artemis/DNA-PKcs arvellaan olevan nukleaasiaktiivisuutta, joka pystyy pilkkomaan irti 5´ ja 3´ DNA-roikkuvat kohdat
- Ligaatio (Ligation)
XRCC4 toimii säätelyelementtinä, joka stabilisoi DNA-ligaasiIV-entsyymiä, stimuloi sen aktiivisuutta ja ohjaa ligaasia DNA-vauriokohtaan omalla helix- rakenteisella tunnistuskohdallaan ja DNA:ta sitovalla kyvyllään.
On tehty lukuisia tutkimuksia jotta voitaisiin identifioida muitakin NHEJ-järjestelmän komponentteja.
http://www.ebi.ac.uk/interpro/potm/2004_7/Page2.htm
KOMMENTTI
Tässä huomataan, että retrovirus pystyy käyttämään solun DNA:n omaa korjausjärjestelmää ja asentaa proviruksensa DNA-muotoisena tällä järjestelmällä isäntäkehon genomiin, eikä järjestelmä tai muu järjestelmä tunnista vierasta DNA:ta. Tällä tavalla HIV-1 virus on tositaiseksi pandeminen kautta maailman ja tuottaa genomin instabiliteetin.
Tässä alla olevassa kuvassa retrovirus toimii miltei kuten proteiini Ku joka lähtee korjaamaan virhettä, mutta retrovirus tekee virheen isäntäsolun DNA:han. ja leikkaa sen poikki ja sitten ”korjaa ”asettamalla oman itsensä väliin ja herättää korjausjärjestelmän isäntäsolun DNA.n katkaisemalla. Siis (ainakin) NHEJ järjestelmä on retroviruksen orjuuttama.
http://www.nature.com/cdd/journal/v12/n1s/images/4401573f1.jpg
http://www.nature.com/cdd/journal/v12/n1s/full/4401573a.html#fig1
2.6.2009 17:16.Päivitys 4.2. 2015.
Prenumerera på:
Inlägg (Atom)