
(Vaikuttaa siltä että IP6K2 /IP7 tehostaa kaspaasi-8 kautta tapahtuvaa apoptoosia.
Tässä kuvaan tavallisen kertomuksen kaspaaseista lähteenä Dionisios Chrysiksen väitöskirjan taustateksti. The Role of apoptosis in growth plate cartilage during normal and abnormal growth. STH 2004. ISBN 91-628-6293-6.)
Kaspaasit osavaikuttajina apoptoosissa
APOPTOOSI on fysiologinen tapahtuma, jossa solu katoaa tulehduksettomasti
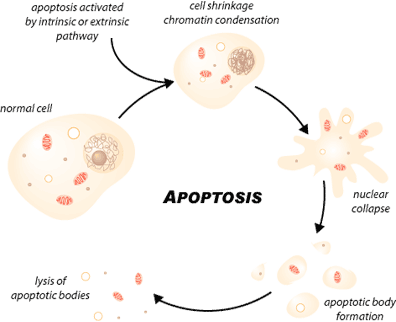
Sellainen solukuolema on ohjelmoitua, missä on erityiset morfologiset ja biokemialliset piirteet, kuten tumakromatiinin tiivistyminen ja DNA:n fragmentoituminen, solun tilavuuden vähenemä ja plasmakalvon muuntumiset, apoptosomien muodostuminen. Näistä seikoista tunnistetaan apoptoosi solun ulkopuolelta käsin ja sellainen solu fagosytoituu. Tämä on fysiologinen prosessi, joka on tärkeä solujen normaalikasvulle ja homeostaasille monisoluisissa elimissä ( Kaikki yhden puolesta ja yksi kaikkien puolesta –periaatteella). Myös apoptoosi on tässä tärkeä geenisäätöinen tekijä yksittäisen orgaanin ja täten koko henkilön kannalta.
Immuunisysteemin säätelyssä tarvitaan apoptoosikykyä.
Tumorogeneesissä on apoptoosilla osuus.
SOLUN GEENISTÖ kantaa apoptoosikykyä.
Miten solu voi apoptoitua?
Se järjestää ohjelman, joka toimii loppuun asti senkin jälkeen, kun solun geenistö on hajautunut. Geneettisesti koodattu ohjelma on evoluutiossa hyvin säilyneenä soluissa ja johtaa tarvittaessa yksityisen solun tehokkaaseen, loogiseen ja siistiin apoptoitumiseen. Solu ikäänkuin ”haihtuu olemattomiin”- Esim vanhetessa keho ikään kuin näkymättömästi kutistuu kaikilta kudoksiltaan fysiologisesti ja loogisesti kuitenkin yksilön elinkyvyn ja tahdonalaisen suuntauksen hyväksi. Tahdonalainen aivotoiminta on korkein määrääjä, arvelen.
APOPTOOTTINEN KASKADI
Apotoosi on jaettavissa kolmeen päävaiheeseen
• Aloitusvaihe, initiaatio
• Exekutiivinen vaihe, lopullinen päätös apoptoitumisesta tehdään.
• Resolutiivinen vaihe, lopputapahtumat.
1. APOPTOOSIN ALKAA jokin seikka, triggeri, kuten kasvutekijän puuttuminen tai vähäisyys, deprivaatio, kuolonreseptoriin sitoutunut ligandi tai genotoksisen tekijän vaikutus. Nämä aiheuttavat, että solu havaitsee elimistölle ja genomille parhaimmaksi ratkaisuksi katoamisen soluyhteiskunnasta. Yksityisen solun genomi asentuu apoptoosiin. Apoptoosin merkitys on integroituna loppusummana kuitenkin globaalin genomin paras mahdollinen elossapysyminen. Apoptoosissa on logiikkaa. Esim. ihminen vanhetessaan pienenee kaikin tavoin ja saattaa täten säilyttää pirteän henkisen struktuurin ja elämänlaadun ja liikkuvuuden ja tarve liikkumiseen /liikunta pitää yllä lokomotorista struktuuria feed backin omaisesti.
2. PÄÄTÖS APOPTOITUMISESTA, Kun exekutiivinen vaihe on tapahtunut, siinä solun genomi alkaa hajota, eikä solu sen jälkeen voi päättää mitään toista muutosta. Spesifiset proteaasit ja nukleaasit hajoittavat rakenteen ja signaloivat proteiinit sekä geneettisen materiaalin johtaen erityisiin morfologisiin muutoksiin.
3. RESOLUUTIOVAIHEESSA solupinta antaa hätämerkkiä lähistöllä oleville terveille soluille, joten ne havaitsevat tämän solun fagosytoosivalmiuden merkit ja niin ne fagosytoivat sen jäänteet. Tämä varmistaa, että mitään itsestään kasvavaa geneettistä materiaalia ei jää jäljelle, sillä geneettinen materiaali on itseään kopioivaa, varsinkin kun se on elävän kehon sisällä, lähellä toisia soluja, jonne se voisi translokoitua. Siksi apoptoosin täydellinen onnistuminen on fysiologisesti ottaen soluyhteiskunnan terveyttä.
MISTÄ TUNNISTAA APOPTOOSIVAIHEITA LÄPIKÄYVÄN SOLUN
Näillä soluilla on morfologisia ja biokemiallisia piirteitä. Nähdään muutos solukalvon integriteetissä, kromatiinia aggregoituu, tuma ja sytoplasma kondensoituu ja tapahtuu sytoplasman ja tuman erkanemista kalvojen ympäröimiksi rakkuloiksi, apoptosomeiksi, joissa on sisällä ribosomeja, morfologisesti ehjiä mitokondrioita ja tumamateriaalia. Solu integroi ja tarkentaa apoptoosinsa asettaen nämä geneettisesti aktiivit arat ainekset erikseen paketoituna odottamaan jatkokäsittelyä. Koeputkessa nämä apoptoosipaketit tunnistetaan nopeasti ja makrofagit alkavat fagosytoida niitä tai lähellä oleva epiteelikin voi toimia fagosytoijana.
Tähän apoptoottisten solujen tehokkaaseen poistamiseen in vivo ei tarvitse aktivoida tulehdusvastetta. Asia hoituu ikäänkuin sopusoinnissa lähisolujen kanssa, missä kaikille on hyödyksi tulehdukseton miljöö.
http://www.scq.ubc.ca/wp-content/uploads/2006/07/Apoptosis.gif
APOPTOOSITIEN KOMPONENTIT
Miten tällainen hieno järjestelmä toimii?
Nisäkäskehossa tapahtuvan apoptoosin kaikkia yksityiskohtia ei ole selvitetty , mutta on voitu todeta yhtäläisyyksiä nematoda-solujen apoptoosiin. Kuolonsäätäjiä on näissä ainakin kaksi pääluokkaa:
• Onkogeenien perhe
• Cysteiiniproteaasien perhe, kaspaasit
ONKOGEENIEN PERHE, Bcl-2
Prototyyppinä on Bcl-2. Ryhmään kuuluu Bcl-2 ja Bax.
ANTIAPOPTOOTTISIA eli kuolon antagonisteja, vastavaikuttajia, ovat Bcl-2, Bcl-x1, Bcl-w, Mcl-1, Bfl-1, Brag.
PROAPOPTOOTTISIA eli kuolon agonisteja, myötävaikuttajia, ovat Bax, Bak, Bcl-xS, Bik.
http://www.ncbi.nlm.nih.gov/bookshelf/picrender.fcgi?book=eurekah∂=A8980&blobname=ch736f2.gif
CYSTEIINIPROTEAASIEN PERHE , Caspases
ICE-Like molecules, caspases
Interleukiini 1-beta (IL-1b) converting enzyme like molecules
Näiden toinen nimi siis on kaspaasit. Kaspaasit ovat apoptoosin effektoreja ja vastuussa proteiinien spesifisten tunnistuskohtien proteolyyttisesta pilkkoamisesta. Pääasiassa ne samalla inaktivoivat kohdeproteiinia. Tosin on muutamia tapauksia, että ne aktivoivat proteiinifunktion.
Mitä proteiineja kaspaasit pilkkovat?
Kaspaasit hajoittavat niitä proteiineja, jotka ovat osallistuneet olennaisina komponentteina sytoplasman ja tuman struktuurien ylläpitämiseen. Tällaisia ovat lamiini A ja fodriini. DNA-korjausjärjestelmän jäsenet ovat niitä myös, kuten PARP ( poly-ADP-riboosi-polymeraasi)
DNA:n hajoamisen estäjät ovat myös niitä: Tällainen on CAD-estäjä, kaspaasiaktivoidun desokxyribonukleaasin estäjä.
Solusyklin säätelyyn osallistuvat proteiinit ovat myös kaspaasien kohteina, kuten esim pRb( retinoblastoomaproteiini)
• Initiaattorikaspaasit (-8, -9)
• Exekutoriset kaspaasit (-3,-6,-7)
Normaalisti kaspaasit ovat inaktiiveina proentsyymeinä läsnä solussa ja ne tarvitsevat sen takia aktivoituakseen proteolyyttisen prosessoitumisen. Aktivoituminen tapahtuu joko autokatalyyttisesti tai toisten kaspaasien vaikutuksesta.
Apoptoosin aloittajakaspaasit, initiaattorit ovat kaspaasi-8 ja kaspaasi-9
Näillä aktivoituja muita kaspaaseja kutsutaan exekutorisiksi kaspaaseiksi kuten kaspaasi-3, kaspaasi-6 ja kaspaasi-7.
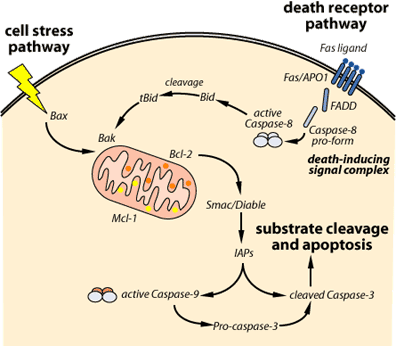
Solun ulkoiset ja sisäiset APOPTOOTTISET TIET
Vaikka apoptoosin alkamisen tarkkoja mekanismeja ei tunnetakaan in vivo, tiedetään, että vasteen kehittyminen on riippuvainen triggeröivästä tekijästä ja on solutyypille spesifinen.
Apoptoosin signaalitransduktio saattanee edetä kahdella toisistaan eriävällä reitillä.
• reseptorivälitteinen eli ulkoinen apoptoositie EXTRINSIC
• mitokondriavälitteinen eli sisäinen apoptoositie, INTRINSIC.
ULKOTIE (EXTRINSIC ) reseptorivälitteinen apoptoositie
Tämä tie vaatii ligandin(L) sitoutumisen kuolonreseptoriin solun pinnalla.
Esimerkiksi TNF( tuumorinekroositekijä) sitoutuu reseptoriin TNF-1R ja rekrytoi kaksi signaalitransduktiomolekyyliä
1. TNF-1R –assosioituneen kuolondomaanin(death domain, DD) proteiinin , TRADD
2. Fas-assosioituneen proteiinin, jolla myös on kuolondomaani, FADD.
TNFR-perheen kuolonreseptoreihin DR kuuluu TNFR1, Fas(CD95), DR3/WSL sekä
TNF-kaltainen apoptoosia indusoiva ligandi TRAIL/Apo21 reseptori (TRAIL-R1/DR4, TRAIL-R2/DR5).
Tämän ryhmän jäsenille on luontaista, että niillä on 2-5 cysteiinirikasta extrasellulaarista toistojaksoa. Kuolonreseptoreilla on myös solunsisäinen DD-domaanin nimellä kutsuttu aminohappojakso C-terminaalissa.
Kun nämä kuolonreseptorit ovat sitoutuneet vastaaviin ligandeihin (TNF tai lymfotoksiini, FasL, DR3-ligandi tai TRAIL/Apo-21), apoptoosi voi käynnistyä.
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah∂=A8980&rendertype=figure&id=A8987
Kaksi prokaspaasi-8-molekyyliä voi prosessoida toisensa aktiiviin muotoon. Aktiivi kaspaasi-8 voi siten triggeröidä päälle apoptoottisen kaspaasikaskadin.
(Uutta tietoa on , että PP—IP5 ja IP6K2 vaikuttavat kaspaasi-8 teitse apoptoosia edistävästi).
SISÄTIE eli mitokondriaalinen apoptoositie
Tieto lisääntyy mitokondrian essentiellistä osuudesta useissa apoptoosimuodoissa, koska se vapauttaa apoptogeenisiä tekijöitä kuten sytokromi C- molekyylin (Cyt C) intermembraanitilasta sytoplasmaan ja täten aktivoi apoptoosin ”downstream execution” tapahtumat. Cyt c irtoama solun energialaitoksesta, mitokondriasta, on kuin mitokondrian hengen poistuminen.
http://www.ncbi.nlm.nih.gov/bookshelf/picrender.fcgi?book=eurekah∂=A32938&blobname=ch174f3.jp
http://www.reading.ac.uk/nitricoxide/intro/apoptosis/mito.htm
Antiapoptoottiset Bcl-2-proteiiniperheen jäsenet koettavat pääasiassa estää mitokondriaalisia muutoksia ja täten mitokondriasta johtuvaa apoptoosia. Nämä antiapoptoottiset Bcl-2 ja Bcl-X1 sijaitsevat mitokondrian ulkokalvossa.
Sitävastoin proapoptoottiset jäsenet sijaitsevat joko sytosolissa tai mitokondrian kalvossa. Ryhmät vastavaikuttavat toisiinsa. Antiapoptoottiset koettavat ehkäistä sytokromi C:n irtoamista ja täten pitkittää solun pysymistä hengissä. Mutta proapoptoottiset aloittavat cyt c-hengitysketjutekijän irrottamisella mitokondriasta. Tämä taas aktivoi executorisia kaspaaseja muodostamalla apoptosomin.
Apoptosomin muodostaa cyt c, proteaasia aktivoiva tekijä-1 Apaf-1 ja prokaspaasi-9.
http://www.reading.ac.uk/nitricoxide/intro/apoptosis/mito2.jpg
On toinenkin mitokondriaalinen kaspaasiaktivaattori.
Tämän nimi on Smac/DIABLO, ja sitä vapautuu mitokondriasta c-sytokromin mukana. Se edistää kaspaasiaktivaatiota liittymällä apoptosomiin ja estämällä apoptoosin inhibiittoriproteiinia IAP.
http://www.rsc.org/ej/NP/2001/a909080k/a909080k-f6.gif
Smac/DIABLO sitoutuu IAP-proteiiniin ja rikkoo sen liittymisen prosessoituvaan kaspaasi-9-molekyyliin ja niin se sallii kaspaasi-9- entsyymin prosessoitua edelleen ja aktivoida kaspaasi-3-proteiinin, joka on executorisia kaspaaseja.
http://www.scq.ubc.ca/wp-content/uploads/2006/07/apoptosispathways.gif
KASPAASEISTA RIIPPUMATTOMAT APOPTOOSITIET:
On osoitusta siitä, että tapahtuu apoptooseja ilman kaspaasien osuutta. Nimittäin vaikka kaspaasit inhiboitaisiin, tapahtuu silti solun kuolemaa ja sen merkkinä tuman kondensaatiota, kalvojen kuplimisia ja solujen kutistumisia. Tästä on päätelty, että kaspaaseja ei tarvita apoptoosin kaikkiin aspekteihin.
AIF
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=eurekah∂=A32938&rendertype=figure&id=A32956
Yksi avainmolekyyli kaspaaseista riippumattomaan solukuolemaan on apoptoosia indusoiva tekijä AIF (Apoptosis Inducing Factor). Solukuolon signaloinnissa AIF translokoituu mitokondriaalisesta intermembraanitilasta tumaan, missä se sitoutuu DNA:han ja aiheuttaa kaspaaseista riippumattoman kromatiinin kondensaation .
5.6.2009 17:39
KOMMENTTINI: IP7 muodon tulee pystyä muuttua IP8 muodoksi, muuten se jää apotoosia stimuloivaksi kun taas IP8 muoto antaa suojaa kromosomille.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Prenumerera på:
Kommentarer till inlägget (Atom)
Inga kommentarer:
Skicka en kommentar