
JC Otto, P Kelly, ST Chiou, JD York Alterations in an inositol phosphate code through synergistic activation of a G protein an Proc Natl Acad Sci U S A (2007) 0:
http://www.gproteins.com/showabstract.php?pmid=17895383
Nisäkkäissä moni solustimulus herättää vasteen G-proteiinin kautta aktivoimalla PLC-entsyymin (fosfolipaasiC) ja se johtaa lipidijohdannasten inositoli 1,4,5-trifosfaattien(IP3) tuotantoon.
Vaikka on hyvin varmistettua, että IP3 voi konvertoitua lukuisiksi inositolifosfaateiksi(IPs) ja pyrofosfaateiksi(PP-IPs) jopa kuuden inositolifosfaattikinaasiluokan (IPKs) kautta, ei olla selvillä siitä, vaikuttaako G-proteiinisignalointi näihin metaboliitteihin.
Tässä artikkelissa tutkijat raportoivat, että Galphaq aktivaatio johtaa vahvaan IP3-stimulaatioon, josta seuraa IP8 muotoon johtava aineenvaihdunta
Jotta saataisin havainnoitua näitä teitä, käytettiin geneettistä häirintää IP-homeostaasiin.
Kun kytketiiniin konstitutionaalisesti aktiivi GalphaqQL ja yksi tai useampi IPK-geenituotteista
ne generoivat synergistisesti dramaattisia muutoksia intrasllulaarisissa IP-välittäjäaine-käyttäytymisissä.
Moni erillinen IP-profiili havainnoitiin eri IPK-kombinaation ilmentymisessä ja siihen kuului muutokset aiemmin aliarvioiduissa IP5 ja IP6-altaissa - niitä oli nimittäin pidetty aiemmin stabiileina metaboliitteina.
Tutkijoitten tiedot linkkivät trimeerisen G-proteiinin aktivaation erittäin runsaaseen metaboliittimäärään IP3:sta eteenpäin ja tarjoaa raamityön jossa oletetaan. että soluilla on koneistoa tuotta IPK.sta riippuva IP-koodi.
Our data link the activation of a trimeric G protein to a plethora of metabolites downstream of IP3 and provide a framework for suggesting that cells possess the machinery to produce an IPK-dependent IP code.
Tutkijat edellyttävät, tosin eivät osoita asiaa, että agonistin indusoimat muutokset sellaisessa koodissa voisivat teoreettisesti ottaen pystyä lisäämään kompleksia signalointia ja sen spesifisyyttä. Organismin kehittymiselle ja solujen adaptaatiolle essentiellit IPK entsyymien tehtävät täsmäävät tutkijoitten hypoteesiin siitä, että on olemassa signaaliteissä relevantti IP koodi.
Kommenttini: Aikoinaan kun pyrin biomedisiiniseen tutkijakouluun täällä Göteborgissa ja sitten sen sijaan pääsin lukemaan dietetiikkaa, halusin tehdä glyserolin aineenvaihdunnasta, sen fosforyloitumisongelmista ja sitemmin fytiinistä (IP6) erään projektin. Mutta nyt näyttää siltä että tuhannet tiedemiehet ovat fytiinin ja kaikkien inositolien koodin IP Cods) kimpussa, joten saan tyytyä vain kirjoitatmaan heidän löytöjään muistiin tähän blogiin. Nobelikin tuli telomeereistä, joita pitää kunnossa osaltaan inositolien fosfaattirikkaasta tuotteesta peräisin oleva uudistusenergia. Fosfaattien aineenvaihdunta on hyvin laaja alue. Fytiinistä ajattelin vain elintarviketekniikan puolista osuutta: elintarviketaulukon tekemistä tulevaista dietääristä, myös fytiinin(inositolin) kattavaa taulukkoa suunnitellen, koska pidän inositolirengasta essentiellinä, ravinnossa tulevana tekijänä.
lördag 12 december 2009
Löytöjä 1995. PACAP ja inositolifosfaatit
DM Simeone, DI Yule, CD Logsdon, JA Williams Ca2+ signaling through secretagogue and growth factor receptors on pancreatic AR42J cells. Regul Pept (1995) 55: 197-206.
Intrasellulaarinen signalointi lisää solunsisäistä kalsiumia:
tämä havaittiin haiman AR42J soluista vasteena agonisteille, joiden reseptori oli G-proteiiniin kytketty: näitä olivat
cholecystokinin (CCK), bombesin, carbachol, substance P, pituitary adenylate cyclase activating peptide (PACAP), bradykinin, ATP, calcitonin gene related peptide (CGRP),
ja vasteena kasvutekijöille EGF ja FGF, joiden reseptorina ovat tyrosiinikinaasit.
Vaste kasvutekijöille oli vahvuudeltaan ja respondoivien solujen määrältä pienempi, mutta riippumaton solunulkoisesta kalsiumista Ca++.
CCK ja karbakoliini vaikuttivat huomattavat nousut inositolifosfaateissa(IPs), mutta niin ei vaikuttanut kasvutekijät.
Fosfolipaasi C-estäjä(PLC-inhibiittori , blokeerasi karbakoliini- ja EGF-vasteet.
Tyrosiinikinaasinestäjä(genesteiini) blokeerasi vasteen EGF:lle, mutta ei vastetta CCK:lle.
Nämä tiedot viitaavat kahdentyyppiseen mekanismiin ARn2J-soluissa.
Sekretagogit vaikuttivat niihin reseptoreihin, jotka olivat kytkeytyneet G-proteiineihin ja indusoivat suuret määrät inositolifosfaatin tuotantoa ja sitä seuraavaa intrasellulaarista kalsiumin mobilisoitumista.
Kasvutekijät vaikuttivat reseptoreihin, jotka signaloivat tyrosiinikinaasin aktiivisuuden välityksellä ja tuottivat tässä solutyypissä rajoitetuin määrin inositolifosfaattia ja pienemmän nousun solunsisäisessä kalsiumissa.
Intrasellulaarinen signalointi lisää solunsisäistä kalsiumia:
tämä havaittiin haiman AR42J soluista vasteena agonisteille, joiden reseptori oli G-proteiiniin kytketty: näitä olivat
cholecystokinin (CCK), bombesin, carbachol, substance P, pituitary adenylate cyclase activating peptide (PACAP), bradykinin, ATP, calcitonin gene related peptide (CGRP),
ja vasteena kasvutekijöille EGF ja FGF, joiden reseptorina ovat tyrosiinikinaasit.
Vaste kasvutekijöille oli vahvuudeltaan ja respondoivien solujen määrältä pienempi, mutta riippumaton solunulkoisesta kalsiumista Ca++.
CCK ja karbakoliini vaikuttivat huomattavat nousut inositolifosfaateissa(IPs), mutta niin ei vaikuttanut kasvutekijät.
Fosfolipaasi C-estäjä(PLC-inhibiittori , blokeerasi karbakoliini- ja EGF-vasteet.
Tyrosiinikinaasinestäjä(genesteiini) blokeerasi vasteen EGF:lle, mutta ei vastetta CCK:lle.
Nämä tiedot viitaavat kahdentyyppiseen mekanismiin ARn2J-soluissa.
Sekretagogit vaikuttivat niihin reseptoreihin, jotka olivat kytkeytyneet G-proteiineihin ja indusoivat suuret määrät inositolifosfaatin tuotantoa ja sitä seuraavaa intrasellulaarista kalsiumin mobilisoitumista.
Kasvutekijät vaikuttivat reseptoreihin, jotka signaloivat tyrosiinikinaasin aktiivisuuden välityksellä ja tuottivat tässä solutyypissä rajoitetuin määrin inositolifosfaattia ja pienemmän nousun solunsisäisessä kalsiumissa.
PACAP, ATP, AMP ja IP8
PACAP signaling
https://www1.qiagen.com/Geneglobe/PathwayView.aspx?pathwayID=341
Ihmisen endokrinologisen järjestelmän korkein adenylsyklaasi(AC) toiminta on PACAP, käpylisäketasossa.
Tämä on kaiken tasapainoisen metabolian avainkarttaa.
Circadinen rytmi, joka ylläpitää kehon oskillatoristen solujen oskillaatioitten alkuvoimaa valon ja pimeyden välisellä aaltoilulla, liittyy tähän järjestelmään ja tuottaa tehokasta aineenvaihduntaa, joka käyttää pimeyttä, lepoa, nukkumisaikaa , yötä, aineenvaihdunnalliseen toipumiseen.
Kun korjataan esim diabeettisuutta kehosta, tulee ottaa huomioon nämä seikat.
Ihmisen hyvinvointi riippuu tämän tason riittävästä energeettisestä tasapainosta.
https://www1.qiagen.com/Geneglobe/PathwayView.aspx?pathwayID=341
Ihmisen endokrinologisen järjestelmän korkein adenylsyklaasi(AC) toiminta on PACAP, käpylisäketasossa.
Tämä on kaiken tasapainoisen metabolian avainkarttaa.
Circadinen rytmi, joka ylläpitää kehon oskillatoristen solujen oskillaatioitten alkuvoimaa valon ja pimeyden välisellä aaltoilulla, liittyy tähän järjestelmään ja tuottaa tehokasta aineenvaihduntaa, joka käyttää pimeyttä, lepoa, nukkumisaikaa , yötä, aineenvaihdunnalliseen toipumiseen.
Kun korjataan esim diabeettisuutta kehosta, tulee ottaa huomioon nämä seikat.
Ihmisen hyvinvointi riippuu tämän tason riittävästä energeettisestä tasapainosta.
Solun energiatilan merkkinä IP8
Solun energiatila heijastuu IP8 tilanteeseen.
Energiatila valvoo bis-difosfoinositolitetrakisfosfaatin synteesiä itsenäisesti ja riippumatta AMP-aktivoidusta proteiinikinaasista
LÄHDE: K. Choi, E. Mollapour, J. H. Choi, S. B. Shears Cellular Energetic Status Supervises the Synthesis of Bis-Diphosphoinositol Tetrakisphosphate Independently of AMP-Activated Protein Kinase Mol. Pharmacol. 2008 74: 527-536.
shears(AT) niehs.nih.gov
ABSTRAKTI
Solut puolustavat aggressiivisesti adenosiininukleotiditasapainoaan; solunsisäiset biosensorit havaitsevat energiatilan vaihteluita ja kommunikoivat toisten soluverkostojen kanssa aloittaakseen sopeuttavia (adaptatiivisia) vasteita.
Tässä artikkelsisa tutkijat osoittavat eräitä uusia elementtejä tästä kommunikaatioprosessista. He osoittavat eräitten suosittujen farmakologisten työvälineitten aiheuttamien bioenergeetisten vaikutusten( alentuneen energiatilan) avulla, että tämä verkkotoiminta kompromisoituu energiatilan mukaan (tässä: energian puutteesta).
Kun soluja stimuloitiin AICAR:illa mikä simuloi kohonneita AMP-pitoisuuksia, aleni IP8 synteesi. IP8 on solunsisäinen signaali( rikasfosfaattinen molekyyli) joka fosforyloi proteiineja kinaaseista( entsyymeistä) riippumattomassa reaktiossa. AICAR vaikutus oli selektiivsesti typistävä. Muitten inositolifosfaattien( köyhempifosfaattisten) pitoisuudet eivät muuttuneet
Sensijaan tiedemiehet tekivät johtopäätöksen, että simuloitaessa adenosiininukleotiditasapainon itsensä heikkenemistä, estyi IP8-syntetisoituminen. Samaan viittaa oligomysiinillä tehdyt tutkimukset, nimittäin se myös kohottaa solun vähäfosfaattisen AMP:n tasoa ja selektiivisesti estää (rikkaan) IP8-synteesin vaikuttamatta muihin IP-muotoihin.
Lisäksi tutkijat osoittivat, että lyhytaikainen IP8-pitoisuuden nousu, mikä tapahtuu normaalisti hyperosmoottisessa stressissä, jäikin heikommaksi eräästä koe-aineesta nimeltä 2-(2-chloro-4-iodo-phenylamino)-N-cyclopropylmethoxy-3,4-difluoro-benzamide (PD184352). Tuota ainetta pidetään tyypillisenä mitogeenistä aktivoidun proteiinikinaasin(MEK) spesifisenä estäjänä. Mutta kun tutkittiinMEK:in vastaista siRNA:ta tai extrasellulaarisen signaalin säätelemää kinaasia havaittiin, että MAPK tie ei ollutkaan mukana asiassa. Sen sijaan tutkijat osoittivat, että IP8 synteesi estyi tuosta koeaineesta sen soluenergiatasapainoon tekemien nonspesifisten vaikutusten takia.
Kaksi muutakin MEK-estäjää tutkittiin ja niista tuli samat vaikutukset ja johtopäätökset. Tästä tutkijat päättelivät, että IP8-molekyylin pitoisuutta ja signalointivoimaa määrää solun adenosiininukleotidien tasapapino, mikä on merkitsevät uusi linkki signaloivien verkostojen ja bioenergeettisten verkostojen kesken.
We conclude that the levels and hence the signaling strength of [PP]2-InsP4 is supervised by cellular adenosine nucleotide balance, signifying a new link between signaling and bioenergetic networks.
Työntaustalla: This research was supported by the Intramural Research Program of the National Institutes of Health/National Institute of Environmental Health Sciences
Termejä:
Adenosiininukleotidejä ovat AMP, ADP, ATP
InsP7, PP-InsP5, diphosphoinositol pentakisphosphate;
[PP]2-InsP4, InsP8, bis-diphosphoinositol tetrakisphosphate;
InsP5, inositol pentakisphosphate;
InsP6, inositol hexakisphosphate;
AICAR, 5-aminoimidazole-4-carboxamide ribonucleoside;
AMPK, AMP-activated protein kinase;
GAPDH, glyceraldehyde 3-phosphate dehydrogenase;
PP-InsP4, diphosphoinositol tetrakisphosphate;
PPIP5K, PP-InsP5 kinase (E.C. 2.7.4.24),
ZMP, 5-amino-4-imidazolecarboxamide riboside monophosphate;
PD184352, 2-(2-chloro-4-iodo-phenylamino)-N-cyclopropylmethoxy-3,4-difluoro-benzamide;
MEK, mitogen-activated protein kinase kinase;
siRNA, small interfering RNA;
ERK, extracellular signal-regulated kinase;
HPLC, high-performance liquid chromatography;
U0126, 1,4-diamino-2,3-dicyano-1,4-bis(methylthio)butadiene;
PD98059, 2′-amino-3′-methoxyflavone.
Received December 21, 2007. Accepted May 6, 2008.
Energiatila valvoo bis-difosfoinositolitetrakisfosfaatin synteesiä itsenäisesti ja riippumatta AMP-aktivoidusta proteiinikinaasista
LÄHDE: K. Choi, E. Mollapour, J. H. Choi, S. B. Shears Cellular Energetic Status Supervises the Synthesis of Bis-Diphosphoinositol Tetrakisphosphate Independently of AMP-Activated Protein Kinase Mol. Pharmacol. 2008 74: 527-536.
shears(AT) niehs.nih.gov
ABSTRAKTI
Solut puolustavat aggressiivisesti adenosiininukleotiditasapainoaan; solunsisäiset biosensorit havaitsevat energiatilan vaihteluita ja kommunikoivat toisten soluverkostojen kanssa aloittaakseen sopeuttavia (adaptatiivisia) vasteita.
Tässä artikkelsisa tutkijat osoittavat eräitä uusia elementtejä tästä kommunikaatioprosessista. He osoittavat eräitten suosittujen farmakologisten työvälineitten aiheuttamien bioenergeetisten vaikutusten( alentuneen energiatilan) avulla, että tämä verkkotoiminta kompromisoituu energiatilan mukaan (tässä: energian puutteesta).
Kun soluja stimuloitiin AICAR:illa mikä simuloi kohonneita AMP-pitoisuuksia, aleni IP8 synteesi. IP8 on solunsisäinen signaali( rikasfosfaattinen molekyyli) joka fosforyloi proteiineja kinaaseista( entsyymeistä) riippumattomassa reaktiossa. AICAR vaikutus oli selektiivsesti typistävä. Muitten inositolifosfaattien( köyhempifosfaattisten) pitoisuudet eivät muuttuneet
Sensijaan tiedemiehet tekivät johtopäätöksen, että simuloitaessa adenosiininukleotiditasapainon itsensä heikkenemistä, estyi IP8-syntetisoituminen. Samaan viittaa oligomysiinillä tehdyt tutkimukset, nimittäin se myös kohottaa solun vähäfosfaattisen AMP:n tasoa ja selektiivisesti estää (rikkaan) IP8-synteesin vaikuttamatta muihin IP-muotoihin.
Lisäksi tutkijat osoittivat, että lyhytaikainen IP8-pitoisuuden nousu, mikä tapahtuu normaalisti hyperosmoottisessa stressissä, jäikin heikommaksi eräästä koe-aineesta nimeltä 2-(2-chloro-4-iodo-phenylamino)-N-cyclopropylmethoxy-3,4-difluoro-benzamide (PD184352). Tuota ainetta pidetään tyypillisenä mitogeenistä aktivoidun proteiinikinaasin(MEK) spesifisenä estäjänä. Mutta kun tutkittiinMEK:in vastaista siRNA:ta tai extrasellulaarisen signaalin säätelemää kinaasia havaittiin, että MAPK tie ei ollutkaan mukana asiassa. Sen sijaan tutkijat osoittivat, että IP8 synteesi estyi tuosta koeaineesta sen soluenergiatasapainoon tekemien nonspesifisten vaikutusten takia.
Kaksi muutakin MEK-estäjää tutkittiin ja niista tuli samat vaikutukset ja johtopäätökset. Tästä tutkijat päättelivät, että IP8-molekyylin pitoisuutta ja signalointivoimaa määrää solun adenosiininukleotidien tasapapino, mikä on merkitsevät uusi linkki signaloivien verkostojen ja bioenergeettisten verkostojen kesken.
We conclude that the levels and hence the signaling strength of [PP]2-InsP4 is supervised by cellular adenosine nucleotide balance, signifying a new link between signaling and bioenergetic networks.
Työntaustalla: This research was supported by the Intramural Research Program of the National Institutes of Health/National Institute of Environmental Health Sciences
Termejä:
Adenosiininukleotidejä ovat AMP, ADP, ATP
InsP7, PP-InsP5, diphosphoinositol pentakisphosphate;
[PP]2-InsP4, InsP8, bis-diphosphoinositol tetrakisphosphate;
InsP5, inositol pentakisphosphate;
InsP6, inositol hexakisphosphate;
AICAR, 5-aminoimidazole-4-carboxamide ribonucleoside;
AMPK, AMP-activated protein kinase;
GAPDH, glyceraldehyde 3-phosphate dehydrogenase;
PP-InsP4, diphosphoinositol tetrakisphosphate;
PPIP5K, PP-InsP5 kinase (E.C. 2.7.4.24),
ZMP, 5-amino-4-imidazolecarboxamide riboside monophosphate;
PD184352, 2-(2-chloro-4-iodo-phenylamino)-N-cyclopropylmethoxy-3,4-difluoro-benzamide;
MEK, mitogen-activated protein kinase kinase;
siRNA, small interfering RNA;
ERK, extracellular signal-regulated kinase;
HPLC, high-performance liquid chromatography;
U0126, 1,4-diamino-2,3-dicyano-1,4-bis(methylthio)butadiene;
PD98059, 2′-amino-3′-methoxyflavone.
Received December 21, 2007. Accepted May 6, 2008.
IP7 tekee nonentsymaattisen fosforylaation proteiineihin
LÄHDE: Science 306 (5704): 2101-2105
Adolfo Saiardi, Rashna Bhandari, Adam C. Resnick, Adele M. Snowman, Solomon H. Snyder
Phosphorylation of Proteins by Inositol Pyrophosphates (2004)
http://stke.sciencemag.org/cgi/content/abstract/sci;306/5704/2101
Inositolipyrofosfaatit IP7 ja IP8 sisältävät korkeaenergisiä pyrofosfaattisidoksia.
The inositol pyrophosphates IP7 and IP8 contain highly energetic pyrophosphate bonds.
"Vaikka niitä on havaittu monissa biologisissa funktioissa niiden molekulaarinen vaikutuskohta/kohdat ei(vät) ole ollut(olleet) selvillä aiemmin. Tässäkin artikkelissa kerrotaan yrityksistä löytää vaikutuskohtia.
Tutkijat käyttivät radioaktiivisella nuklidilla merkattua IP7 molekyyliä ja löysivät monien eukaryoottisten proteiinien fosforylaatioita.
He havaitsivat myös hiivan endogeenisen IP7 fosforyloivan endogeenisia proteiineja. Tämä IP7:n tekemä fosforylaatio on luonteeltaan nonentsymaattinen tapahtuma ja saattanee edustaa uutta (novel) intrasellulaarista signalointimekanismia."
(Kommenttini: Siis "energiakylpy" proteiinille, soveltuu varmaan hyvin suojaamaan kromosomaalista rakennetta ja korjaamaan DNA:ta tai vahvistamaan executiivisia tapahtumia) .
Adolfo Saiardi, Rashna Bhandari, Adam C. Resnick, Adele M. Snowman, Solomon H. Snyder
Phosphorylation of Proteins by Inositol Pyrophosphates (2004)
http://stke.sciencemag.org/cgi/content/abstract/sci;306/5704/2101
Inositolipyrofosfaatit IP7 ja IP8 sisältävät korkeaenergisiä pyrofosfaattisidoksia.
The inositol pyrophosphates IP7 and IP8 contain highly energetic pyrophosphate bonds.
"Vaikka niitä on havaittu monissa biologisissa funktioissa niiden molekulaarinen vaikutuskohta/kohdat ei(vät) ole ollut(olleet) selvillä aiemmin. Tässäkin artikkelissa kerrotaan yrityksistä löytää vaikutuskohtia.
Tutkijat käyttivät radioaktiivisella nuklidilla merkattua IP7 molekyyliä ja löysivät monien eukaryoottisten proteiinien fosforylaatioita.
He havaitsivat myös hiivan endogeenisen IP7 fosforyloivan endogeenisia proteiineja. Tämä IP7:n tekemä fosforylaatio on luonteeltaan nonentsymaattinen tapahtuma ja saattanee edustaa uutta (novel) intrasellulaarista signalointimekanismia."
(Kommenttini: Siis "energiakylpy" proteiinille, soveltuu varmaan hyvin suojaamaan kromosomaalista rakennetta ja korjaamaan DNA:ta tai vahvistamaan executiivisia tapahtumia) .
fredag 11 december 2009
IP7 muotoon IP8 ja vip entsyymit
Hakusana vip-1 and vip-2 enzymes antaa Google hakuna 2730 vastausta. Asia näyttää olevan eläinkoetasossa. Koetan löytää tältä vuodelta uutta mtuta saan uusimpana allaolevaa.
Cloning and Characterization of Two Human VIP1-like Inositol Hexakisphosphate and Diphosphoinositol Pentakisphosphate Kinases. Otan tässä toistamiseen artikkelin
Peter C. Fridy, James C. Otto, D. Eric Dollins, John D. York1
Departments of Pharmacology and Cancer Biology and of Biochemistry, Howard Hughes Medical Institute, Duke University Medical Center, Durham, North Carolina, 27710
To whom correspondence should be addressed: Duke University Medical Center, DUMC Box 3813, Duke University Medical Center, Durham, NC 27710.
Abstrakti. Suomennosta (kesken)
Artikkeli kertoo ihmisten kahden VIP1-kaltaisen kinaasin, IP6-kinaasin (IP6K) ja difosfoinositoli -IP5-kinaasin (IP7K), kloonaamisesta ja luonnehtimisesta.
EUKARYOOTEILLA on lukuisia inositolifosfaatteja(IP) ja difosfoinositolifosfaatteja(PP-IPs), joita myös kutsutaan inositolipyrofosfaateiksi ja ne vaikuttavat kemiallisina koodeina, jotka ovat tärkeitä solunsisäisissä signaaliteissä. IP- ja PP-IP-molekyylijoukkoa tuottaa useat evolutionaalisesti konservoidut IP-kinaasit.
Tässä tutkijat kertovat, miten he saivat luonnehdittua IP6-kinaasin (IP6K) ja IP7-kinaasin (IP7K= diphosphoinositol pentakisphosphate eli PP-IP5 kinase), jotka olivat hiivassa sen Vip1- entsyymin kaltaisia ja ne identifioitiin IP6/IP7 kinaasina.
Koeputkessa VIP1 osoittaa IP6-kinaasi - ja IP7-kinaasiaktiivisuuta ja saa aikaan IP7 synteesin. VIP1 geenin ilmenemä ihmissolussa tuotti korkeat pitoisuudet IP7 , josta taas johtui dramaattinen lisääntymä IP8-muodossa (= bisdiphosphoinositol tetrakisphosphate (PP2-IP4 or IP8).
Missä ihmiskehon kohdassa ilmenee VIP1 geeniä?
Sitä on monissa eri kudoksissa ja erityisen rikkaana tahdonalaisessa lihaksessa, sydämessä ja aivoissa. Jos VIP1 merkattiin ja katsottiin, missä sitä on, niin sitä löydettiin solun sytoplasman non-membraanisessa tilassa, kohdissa missä ei ole kalvorakenteita.
Tutkijat luonnehtivat myös ihmisen ja hiiren VIP2-entsyymin, joka on toisen geenin tuote, mutta 90 %:sti samanlainen kuin VIP1-entsyymi kinaasidomaaniltaan ja se osoitti myös sekä IP6K ja IP7K aktiivisuutta.
Tutkijat päättelivät, että ihmisen VIP1 ja VIP2 toimivat IP6- ja IP7-kinaaseina, jotka vaikuttavat yhteistyössä IP6K/Kcs1-luokan kinaasien kanssa konvertoimassa IP6-muotoa nisäkässoluissa IP8-muotoon. Tämän prosessin on todettu tapahtuvan vasteena erilaisille stimuluksille ja signalointitapahtumille.
PLC:n osuus
INOSITOLIFOSFAATIT (IPs) on sekalainen ryhmä regulatorisia molekyylejä ja ne osallistuvat monenlaisiin solunsisäisiin( itnrasellulaarisiin) signaaliteihin. Solujen stimuloituminen johtaa IPs tuotantoon siten että fosfoinositidit(PIs) alkavat pilkkoutua fosfoinositideille spesifisellä fosfolipaasilla C(1-3), PLC.
Monet muut IPs molekyylit kuten tetrakisfosfaatit(IP4), inositolipentakisfosfaatti(IP5) ja inositolihexacisfosfaatti(IP6) syntyvät useitten evolutionaalisesti konservoituneitten IPKs(2,-4-7) eli inositolifosfokinaasien luokkaan kuuluvien entsyymien perättäisten toimintojen kautta .
Näitten kinaasien geneettisten ja biokemiallisten tutkimusten seurauksena on kyetty linkitsemään muodostuneet IP-tuotteet sellaisiin prosesseihin kuten
Inositolipyrofosfaatit(PP-IPs) raportoitiin ja luonnehdittiin aluksi Dictyostelium discoideumista ja nisäkässoluista ja ne eroavat toisistaan inositolirenkaassa sijaitsevien yhden tai useamman pyrofosfaattiryhmän suhteen.
Evolutionaalisesti konservoituja kinaaseja on havaittu kahta ryhmää tuottamassa näitä inositoli-pyrofosfaattimuotoja.
(I)
IP6K, IP6-kinaasiluokka (Hiivassa Kcs1. Nisäksäsoluissa IHPK1, IHPK2, IHPK3):
Nämä konvertoivat IP6-muodon ( inositolihexacisfosfaattia eli fytiiniä) difosfoinositolipentakisfosfaatiksi PP-IP5. Sen nimenä on myös IP7.
Entsyymi voi myös fosforyloida IP5 muotoa ja tuottaa PP-IP4; IP6.
JOS silmuilevasta hiivasta puuttuu tämä Kcs1, seuraa puutteita vasteessa osmoottiseen stressiin, telomeerien pituuden säädössä , vakuolaarisessa biogeneesissä, endosytoosissa ja muissa soluprossesseissa.
PP-IP5 muoto ainakin koeputkessa toimii fosfaatin luovuttajana ja pystyy fosforyloimaan proteiineja suoraan ilman entsyymin tarvetta. Kuitenkaan ei ole todisteita, jos tämä tapahtuu myös solumiljöössä.
Tulokset viittaavat myös siihen, että IP6-kinaasiaktiivisuus on osallistumassa olennaisen tärkeän bis-difosfo-inositoli-tetra-kis-fosfaatin ( PP-IP4-PP), siis IP8- muodon (PP2-IP4) synteesiin. Se on vahvasti fosforyloitu ja sisältää kaksi pyrofosfaattiryhmää.
Tutkimukset osoittavat, että PP-IP4 tasot vaihtelevat vasteena osmoottiseen tai kuumastressiin sekä hiivassa että nisäkässolussa ja IP8-tasoa saattaa osaltaan säädellä myös MAP-kinaasitie.
IP8 (eli PP2-IP4) ja IP7 ( eli PP-IP-5) näyttävät ilmeisesti omaavan osuutta cAMP-välitteisissä signaalitapahtumissa ja niihin kuuluu myös chemotaxis D. discoideum organismissa; näiden metaboliittien pitoisuuksissa nimittäin tapahtuu merkitseviä muutoksia cAMP-signaloinnin aikana.
(II)
Toinen IP6/IP7-kinaasiluokka on myös löydetty silmuilevasta hiivasta ja sitä merkataan Vip1 nimellä. Tämä aktiivisuus identifioitiin inositolipyrofosfaattisyntaasina IPS1 (=inositol pyrophosphate synthase IPS1) perustuen tietoon vastamuodostuneen PP-IP5 molekyylin akkumuloitumisesta mutanttihiiressä, josta puuttuu sekä Kcs1/IP6K aktiivisuus että myös difosfoinositolifosfaattifosfataasi Dsp1 ( (kcs1Δddp1Δ) aktiivisuus.
Kun puhdistettiin tällaisesta 2-kertaisesti poistogeenisestä hiivakannasta biokemiallisesti puhdasta IP6-kinaasia, saatiin kloonattua Vip1. Ennenkuin keksittiin , että Vip1 omasi sisäsyntyistä IP6-kinaasiaktiivisuutta, oli havaittu, että se voi toimia myös aktiinin polymerisaation ja sytoskeletaalisen funktion säätelijänä.
(Prior to the discovery that Vip1 possessed intrinsic IP6 kinase activity, its Schizosaccharomyces pombe ortholog Asp1 was found to exhibit genetic interactions with actin-related proteins 2/3 (Arp2/3), indicating that it may function as a regulator of actin polymerization and cytoskeletal function).
Oli demonstroitu, että IP6-kinaasi aktiivisuutta tarvittiin tiettyyn geneettiseen interaktioon ja normaaliin solumorfologiaan.
Lisäksi rekombinantin Vip1:n ja Kcs1/IP6K:n fosforiNMR-analyyseillä osoitettiin, että tuottuu toisistaan erotettavissa olevia PP-IP5-lajeja (distinct PP-IP5 species ).
Ottaen huomioon Vip1 entsyymin ja Kcs1/IP6K entsyymien ainutlaatuiset aktiivisuudet osoitettiin, että molemmat entsyymit toimivat myös PP-IP5-kinaaseina ja yhdessä kykenevät fosforyloimaan IP6-muotoa IP8-muotoon( PP2-IP4).
Tässä katsauksessaan tutkijat raportoivat tunnistaneensa, kloonanneensa ja luonnehtineensa kaksi ihmisen Vip1-kaltaista geenituotetta. Heidän tietonsa viitaavat siihen että ihmisen(human) VIP-1 ja VIP2 toimivat molemmat IP6- ja PP-IP5-kinaaseina, joiden tuotteet omaavat osaa laajassa kirjossa erilaisia signaalinjohtoteitä. Tämä työ on merkitsevä edistysaskel laajentamassa käsitystä inositolipyrofosfaattien osuudesta signaloinnissa.
Cloning and Characterization of Two Human VIP1-like Inositol Hexakisphosphate and Diphosphoinositol Pentakisphosphate Kinases. Otan tässä toistamiseen artikkelin
Peter C. Fridy, James C. Otto, D. Eric Dollins, John D. York1
Departments of Pharmacology and Cancer Biology and of Biochemistry, Howard Hughes Medical Institute, Duke University Medical Center, Durham, North Carolina, 27710
To whom correspondence should be addressed: Duke University Medical Center, DUMC Box 3813, Duke University Medical Center, Durham, NC 27710.
Abstrakti. Suomennosta (kesken)
Artikkeli kertoo ihmisten kahden VIP1-kaltaisen kinaasin, IP6-kinaasin (IP6K) ja difosfoinositoli -IP5-kinaasin (IP7K), kloonaamisesta ja luonnehtimisesta.
EUKARYOOTEILLA on lukuisia inositolifosfaatteja(IP) ja difosfoinositolifosfaatteja(PP-IPs), joita myös kutsutaan inositolipyrofosfaateiksi ja ne vaikuttavat kemiallisina koodeina, jotka ovat tärkeitä solunsisäisissä signaaliteissä. IP- ja PP-IP-molekyylijoukkoa tuottaa useat evolutionaalisesti konservoidut IP-kinaasit.
Tässä tutkijat kertovat, miten he saivat luonnehdittua IP6-kinaasin (IP6K) ja IP7-kinaasin (IP7K= diphosphoinositol pentakisphosphate eli PP-IP5 kinase), jotka olivat hiivassa sen Vip1- entsyymin kaltaisia ja ne identifioitiin IP6/IP7 kinaasina.
Koeputkessa VIP1 osoittaa IP6-kinaasi - ja IP7-kinaasiaktiivisuuta ja saa aikaan IP7 synteesin. VIP1 geenin ilmenemä ihmissolussa tuotti korkeat pitoisuudet IP7 , josta taas johtui dramaattinen lisääntymä IP8-muodossa (= bisdiphosphoinositol tetrakisphosphate (PP2-IP4 or IP8).
Missä ihmiskehon kohdassa ilmenee VIP1 geeniä?
Sitä on monissa eri kudoksissa ja erityisen rikkaana tahdonalaisessa lihaksessa, sydämessä ja aivoissa. Jos VIP1 merkattiin ja katsottiin, missä sitä on, niin sitä löydettiin solun sytoplasman non-membraanisessa tilassa, kohdissa missä ei ole kalvorakenteita.
Tutkijat luonnehtivat myös ihmisen ja hiiren VIP2-entsyymin, joka on toisen geenin tuote, mutta 90 %:sti samanlainen kuin VIP1-entsyymi kinaasidomaaniltaan ja se osoitti myös sekä IP6K ja IP7K aktiivisuutta.
Tutkijat päättelivät, että ihmisen VIP1 ja VIP2 toimivat IP6- ja IP7-kinaaseina, jotka vaikuttavat yhteistyössä IP6K/Kcs1-luokan kinaasien kanssa konvertoimassa IP6-muotoa nisäkässoluissa IP8-muotoon. Tämän prosessin on todettu tapahtuvan vasteena erilaisille stimuluksille ja signalointitapahtumille.
PLC:n osuus
INOSITOLIFOSFAATIT (IPs) on sekalainen ryhmä regulatorisia molekyylejä ja ne osallistuvat monenlaisiin solunsisäisiin( itnrasellulaarisiin) signaaliteihin. Solujen stimuloituminen johtaa IPs tuotantoon siten että fosfoinositidit(PIs) alkavat pilkkoutua fosfoinositideille spesifisellä fosfolipaasilla C(1-3), PLC.
Monet muut IPs molekyylit kuten tetrakisfosfaatit(IP4), inositolipentakisfosfaatti(IP5) ja inositolihexacisfosfaatti(IP6) syntyvät useitten evolutionaalisesti konservoituneitten IPKs(2,-4-7) eli inositolifosfokinaasien luokkaan kuuluvien entsyymien perättäisten toimintojen kautta .
Näitten kinaasien geneettisten ja biokemiallisten tutkimusten seurauksena on kyetty linkitsemään muodostuneet IP-tuotteet sellaisiin prosesseihin kuten
- metallien kelatoiminen
- jonikanavien säätely
- transkriptio
- kromatiinin uudelleenmuokkaus (chromatin remodelling)
- nukleaarisen mRNA:n uloskuljettaminen(nuclear mRNA export)
- apoptoosi
- RNA:n editointi (RNA editing)
- fosfaattiherkistys (phosphate sensing)
- auksiinisignalointi (auxin signaling)
- asianmukainen elinten kehitys (proper organism development)
Inositolipyrofosfaatit(PP-IPs) raportoitiin ja luonnehdittiin aluksi Dictyostelium discoideumista ja nisäkässoluista ja ne eroavat toisistaan inositolirenkaassa sijaitsevien yhden tai useamman pyrofosfaattiryhmän suhteen.
Evolutionaalisesti konservoituja kinaaseja on havaittu kahta ryhmää tuottamassa näitä inositoli-pyrofosfaattimuotoja.
(I)
IP6K, IP6-kinaasiluokka (Hiivassa Kcs1. Nisäksäsoluissa IHPK1, IHPK2, IHPK3):
Nämä konvertoivat IP6-muodon ( inositolihexacisfosfaattia eli fytiiniä) difosfoinositolipentakisfosfaatiksi PP-IP5. Sen nimenä on myös IP7.
Entsyymi voi myös fosforyloida IP5 muotoa ja tuottaa PP-IP4; IP6.
JOS silmuilevasta hiivasta puuttuu tämä Kcs1, seuraa puutteita vasteessa osmoottiseen stressiin, telomeerien pituuden säädössä , vakuolaarisessa biogeneesissä, endosytoosissa ja muissa soluprossesseissa.
PP-IP5 muoto ainakin koeputkessa toimii fosfaatin luovuttajana ja pystyy fosforyloimaan proteiineja suoraan ilman entsyymin tarvetta. Kuitenkaan ei ole todisteita, jos tämä tapahtuu myös solumiljöössä.
Tulokset viittaavat myös siihen, että IP6-kinaasiaktiivisuus on osallistumassa olennaisen tärkeän bis-difosfo-inositoli-tetra-kis-fosfaatin ( PP-IP4-PP), siis IP8- muodon (PP2-IP4) synteesiin. Se on vahvasti fosforyloitu ja sisältää kaksi pyrofosfaattiryhmää.
Tutkimukset osoittavat, että PP-IP4 tasot vaihtelevat vasteena osmoottiseen tai kuumastressiin sekä hiivassa että nisäkässolussa ja IP8-tasoa saattaa osaltaan säädellä myös MAP-kinaasitie.
IP8 (eli PP2-IP4) ja IP7 ( eli PP-IP-5) näyttävät ilmeisesti omaavan osuutta cAMP-välitteisissä signaalitapahtumissa ja niihin kuuluu myös chemotaxis D. discoideum organismissa; näiden metaboliittien pitoisuuksissa nimittäin tapahtuu merkitseviä muutoksia cAMP-signaloinnin aikana.
(II)
Toinen IP6/IP7-kinaasiluokka on myös löydetty silmuilevasta hiivasta ja sitä merkataan Vip1 nimellä. Tämä aktiivisuus identifioitiin inositolipyrofosfaattisyntaasina IPS1 (=inositol pyrophosphate synthase IPS1) perustuen tietoon vastamuodostuneen PP-IP5 molekyylin akkumuloitumisesta mutanttihiiressä, josta puuttuu sekä Kcs1/IP6K aktiivisuus että myös difosfoinositolifosfaattifosfataasi Dsp1 ( (kcs1Δddp1Δ) aktiivisuus.
Kun puhdistettiin tällaisesta 2-kertaisesti poistogeenisestä hiivakannasta biokemiallisesti puhdasta IP6-kinaasia, saatiin kloonattua Vip1. Ennenkuin keksittiin , että Vip1 omasi sisäsyntyistä IP6-kinaasiaktiivisuutta, oli havaittu, että se voi toimia myös aktiinin polymerisaation ja sytoskeletaalisen funktion säätelijänä.
(Prior to the discovery that Vip1 possessed intrinsic IP6 kinase activity, its Schizosaccharomyces pombe ortholog Asp1 was found to exhibit genetic interactions with actin-related proteins 2/3 (Arp2/3), indicating that it may function as a regulator of actin polymerization and cytoskeletal function).
Oli demonstroitu, että IP6-kinaasi aktiivisuutta tarvittiin tiettyyn geneettiseen interaktioon ja normaaliin solumorfologiaan.
Lisäksi rekombinantin Vip1:n ja Kcs1/IP6K:n fosforiNMR-analyyseillä osoitettiin, että tuottuu toisistaan erotettavissa olevia PP-IP5-lajeja (distinct PP-IP5 species ).
Ottaen huomioon Vip1 entsyymin ja Kcs1/IP6K entsyymien ainutlaatuiset aktiivisuudet osoitettiin, että molemmat entsyymit toimivat myös PP-IP5-kinaaseina ja yhdessä kykenevät fosforyloimaan IP6-muotoa IP8-muotoon( PP2-IP4).
Tässä katsauksessaan tutkijat raportoivat tunnistaneensa, kloonanneensa ja luonnehtineensa kaksi ihmisen Vip1-kaltaista geenituotetta. Heidän tietonsa viitaavat siihen että ihmisen(human) VIP-1 ja VIP2 toimivat molemmat IP6- ja PP-IP5-kinaaseina, joiden tuotteet omaavat osaa laajassa kirjossa erilaisia signaalinjohtoteitä. Tämä työ on merkitsevä edistysaskel laajentamassa käsitystä inositolipyrofosfaattien osuudesta signaloinnissa.
IPs-signaloinnin ja chekpointkinaasiperheen linkkiytyminen
Inositolifisfosfaattisignalointi säätelee telomeerien pituutta.
York SJ, Armbruster BN et al. Inositol diphosphate signaling regulates telomere length. J Biol Chem. 2005 Feb 11;280(6):4264-9. Epub 2004 Nov 23.PMID: 15561716
PLC-entsyymistä riippuva inositolifosfaattisignalointitie saa aikaan erilaisia välittäviä signaaleita (messengers), joita muodostuu IP3:sta käsin ja ne kontrolloivat geeniexpressiota ja mRNA:n -kuljetusta ulos tumasta.
PLC =phospholipase C
IP3 on tässä inositol 1,4,5-trisphosphate.
Tässä artikkelissa tutkijat raportoivat telomeerien pituuden säätelyä PP-IP4 molekyylillä, jota tuottaa KCS1 geenituote.
PP-IP4 = diphosphorylinositol tetrakisphosphate.
Jos PP-IP4 tuottuminen katoaa, telomeerit alkavat muodostua pitemmiksi.
Jos PP-IP4 molekyyliä tuottuu ylimäärin, telomeerit lyhenevät.
Tämä vaikutus vaatii Tel1 läsnäoloa( hiivalla tämä vastaa ihmisen ATM (= protein mutated in the human disease ataxia telangiectasia).
Nämä tiedot osoittavat, että on in vivo olemassa säätelylinkki inositolipolyfosfaattisignaloinnin ja checkpoint kinaasien perheen kesken ja samalla kuvaavat kolmannen nukleaarisen prosessin, jota moduloi PLC aktivaatio.
York SJ, Armbruster BN et al. Inositol diphosphate signaling regulates telomere length. J Biol Chem. 2005 Feb 11;280(6):4264-9. Epub 2004 Nov 23.PMID: 15561716
PLC-entsyymistä riippuva inositolifosfaattisignalointitie saa aikaan erilaisia välittäviä signaaleita (messengers), joita muodostuu IP3:sta käsin ja ne kontrolloivat geeniexpressiota ja mRNA:n -kuljetusta ulos tumasta.
PLC =phospholipase C
IP3 on tässä inositol 1,4,5-trisphosphate.
Tässä artikkelissa tutkijat raportoivat telomeerien pituuden säätelyä PP-IP4 molekyylillä, jota tuottaa KCS1 geenituote.
PP-IP4 = diphosphorylinositol tetrakisphosphate.
Jos PP-IP4 tuottuminen katoaa, telomeerit alkavat muodostua pitemmiksi.
Jos PP-IP4 molekyyliä tuottuu ylimäärin, telomeerit lyhenevät.
Tämä vaikutus vaatii Tel1 läsnäoloa( hiivalla tämä vastaa ihmisen ATM (= protein mutated in the human disease ataxia telangiectasia).
Nämä tiedot osoittavat, että on in vivo olemassa säätelylinkki inositolipolyfosfaattisignaloinnin ja checkpoint kinaasien perheen kesken ja samalla kuvaavat kolmannen nukleaarisen prosessin, jota moduloi PLC aktivaatio.
IP2K on inositoli 6-/3-/5- kinaasi, IPx multikinaasi
4. Hiivasolussa on tehty (2005) tutkimus Ins-(1,4,5)-P3 inositolitrifosfaatin aloittaman (uuden) inositolipolyfostaattitien molekulaariseksi määrittämiseksi.
Seeds AM, Bastidas RJ, York JD. Molecular definition of a novel inositol polyphosphate (IPx) metabolic pathway initiated by inositol 1,4,5-trisphosphate 3-kinase activity in Saccharomyces cerevisiae. J Biol Chem. 2005 Jul 29;280(30):27654-61. Epub 2005 Jun 8.PMID: 15944147
Inositolipolyfosfaatteja (IPx) ja inositolipyrofosfaatteja (PP-IPx) syntetisoituu inositolitrifosfaatista IP3 käsin, mutta on ajateltu, että IP3 molekyylin täytyy olla silloin muodossa inositoli1,4,5-trifosfaatti ( eli I(1,4,5)P3), jota näitä poly-muotoja voi kehittyä.
Tähän alkumuokkaukseen tarvitaan IPK2 kinaasiaktiivisuudet: eri hiiliaseman fosforyloijia, 6-/3-/5-kinase.
Tämä kinaasi on tunnettu myös nimellä Arg82 tai sitten inositolipolyfosfaatti- multikinaasina eli nimellä inositol polyphosphate multikinase).
Tässä työssään tutkijat katsoivat IP-aineenvaihdunnassa esiintyviä eroja mainittujen 6- ja 3- kinaasien kesken (distinct roles for I(1,4,5)P3 6- versus 3-kinase activities in IP metabolism and cellular functions reported for Ipk2).
3-kinaasi aloitti uuden synteesitien , josta tuli yli 11 kpl molekyylejä IPs ja PP-IPs.
Primääri synteesitie oli I(1,4,5)P3 --> I(1,3,4,5)P4 --> I(1,2,3,4,5)P5 --> PP-IP4 --> PP2-IP3 ja vaati Kcs1 (tai mahdollisesti Ipk2).
Tutkijat tekivät päättelyn myös uuden IP3-kinaasin olemassaolosta, jolla on merkitystä nukleosytoplasmisissa prosesseissa ja se vaikuttaa IPs ja PP-IPs molekyylimäärien keskinäisiin kompensaatioihin ( toisiaan korvaavaisuuteen).
Seeds AM, Bastidas RJ, York JD. Molecular definition of a novel inositol polyphosphate (IPx) metabolic pathway initiated by inositol 1,4,5-trisphosphate 3-kinase activity in Saccharomyces cerevisiae. J Biol Chem. 2005 Jul 29;280(30):27654-61. Epub 2005 Jun 8.PMID: 15944147
Inositolipolyfosfaatteja (IPx) ja inositolipyrofosfaatteja (PP-IPx) syntetisoituu inositolitrifosfaatista IP3 käsin, mutta on ajateltu, että IP3 molekyylin täytyy olla silloin muodossa inositoli1,4,5-trifosfaatti ( eli I(1,4,5)P3), jota näitä poly-muotoja voi kehittyä.
Tähän alkumuokkaukseen tarvitaan IPK2 kinaasiaktiivisuudet: eri hiiliaseman fosforyloijia, 6-/3-/5-kinase.
Tämä kinaasi on tunnettu myös nimellä Arg82 tai sitten inositolipolyfosfaatti- multikinaasina eli nimellä inositol polyphosphate multikinase).
Tässä työssään tutkijat katsoivat IP-aineenvaihdunnassa esiintyviä eroja mainittujen 6- ja 3- kinaasien kesken (distinct roles for I(1,4,5)P3 6- versus 3-kinase activities in IP metabolism and cellular functions reported for Ipk2).
3-kinaasi aloitti uuden synteesitien , josta tuli yli 11 kpl molekyylejä IPs ja PP-IPs.
Primääri synteesitie oli I(1,4,5)P3 --> I(1,3,4,5)P4 --> I(1,2,3,4,5)P5 --> PP-IP4 --> PP2-IP3 ja vaati Kcs1 (tai mahdollisesti Ipk2).
Tutkijat tekivät päättelyn myös uuden IP3-kinaasin olemassaolosta, jolla on merkitystä nukleosytoplasmisissa prosesseissa ja se vaikuttaa IPs ja PP-IPs molekyylimäärien keskinäisiin kompensaatioihin ( toisiaan korvaavaisuuteen).
Inositolipyrofostaatti on pääsyt Karolinskaan
Löysin Google haulla tutkimustyön, jota tehdään Karoliinisen instituutin helmoissa ja siihen kuuluu myös osa, jossa –sivumennen- katsotaan inositolipyrofosfaattien osuutta betasolujen signalointiin. Haa. Onnea vaan. On vain niin, että inositoliaineenvaihdunta tullee varmasti nielemään vähitellen tutkijan koko energian. Se ei ole niin valmiina, että sitä voi sivumennen tutkia.
Tässä allaolevassa kuvassa on jo tunnettu 3-kinaasi mainittu kuvassa, se kinaasi, jota caffeiini estää ja on hyödyksi antidiabeettisella tavalla.
http://switchgeargenomics.com/wp-content/uploads/2009/08/insulin-receptor-pic.jpg
Meneillään oleva tutkimustyö:
Berggren, Per-Olof. Signal-transduction in the pancreatic beta cell.
Karolinska Institutet - Institutionen för molekulär medicin och kirurgi. 2008-11-04
Suomennan tutkijan projektin alustusta.
(1) Työn tarkoituksena on saada ymmärrystä niistä solumekanismeista, jotka säätelevät insuliinin erittymistä beeta-soluista normaalioloissa ja miksi mekanismit eivät toimi diabeteksessa.
(2) Tässä yhteydessä on eräitä tärkeitä asioita selvitettävänä.
Ensinnäkin täytyy ymmärtää
· ne signaalit, joita verensokeri kehkeyttää
· sekä ne eri aineet, joita erittyy muista kuin beetasoluista haiman hormoneita vapauttavasta osasta ja ympäröivistä hermoista
· ja myös ne aineet, jotka pääsevät beetasoluun verisuonisysteemin kautta.
(3) Näiden signaalien täytyy tulla beetasolun integroimiksi, jotta syntyisi ne edellytykset, mitä tarvitaan insuliinin muodostumiseen ja sen vapauttamiseen.
(4) Toiseksi on käsitettävä niitä solumekanismeja, jotka aktivoituvat, kun beetasolu in integroimassa eri signaaleita.
(5) Kolmanneksi on ymmärrettävä, miten beetasolu toimii elävässä organismissa, kun haimarauhasen hormoneja erittävästa osasta on huolehtimassa sekä verisuonet että hermot.
(6) Tämän takia tutkijan mielenkiinnon kohteena on seuraavat seikat.
1. Solufunktiolle spesifiset ja tärkeät valkuaisaineet, proteiinit tai jonikanavat, jotka kuljettavat varauksellisia Ca++ atomeita (kalsiumjoneja) beeta-soluun ja miten nämä käyttäytyvät normaaliolosuhteissa ja diabeteksessa.
Hur kalciumjonsignalen beter sig och vad detta betyder för beta cellens funktion och överlevnad.
3. Kuinka varsinainen insuliinia vapauttava koneisto vaikuttuu, jos manipuloidaan joittenkin avaintekijöitten saatavuutta.
4. Kuinka muodostuu eräät spesifiset aineet, joita sanotaan inositolipyrofosfaateiksi ja mikä merkitys näillä on beetasolujen funktiolle ja elossapysymiselle.
2009-12-11 11:26
(Kommenttini: Tutkija on oikeilla jäljillä. Toivon menestystä!)
Betaolun funktiossakin inositoliaineenvaihdunta on kuin Spanska trappan Roomassa. Kun niitä aikansa nousee, pääsee ylätasanteelle (= pyrofosfaattien integroitu säätely), jossa on Villa Medici ja valtavan suuri paratiisin puu) ja saa hyvät näköalat asiaan- Roomaan... tarkoitan, että kun joka rappu onnistuu jotenkuten, niin että tulee lopulta ylätasanteelle, diabeettista solua ei ole, vaan keho toimii normaalisti, sen diabeettisuus on ylitetty ongelma.
Mutta inositolijärjestelmän jokainen molekyyli ja entsyymi on yksi niistä rapuista, ja jos nyt vielä hokee kerran: Pitäisi tehdä elintarviketaulukot fytiinistä ja inositolista, niin tulevaisuus helpottuisi terapiamenetelmien suhteen sitten aikanaan. Inositoli kilpailee sokeriaineenvaihdunnan kanssa jotenkin ja sen exogeenisesti saadun määrän osuus on tuntematon tekijä.
On tuntematonta miten paljon nykyihminen ja entisajan ihminen käyttää/ käytti fytiiniä ravinnossa ja täten saa/ sai niitä valmiita inositolirenkaita kehoon.(entisajan ihmisillä ei ollut inositoliaineenvaihdunnan rasitteena teollsita glukoosi ja muita vastaavia, jotka kilpailevat inositolin kanssa soluliikenteessä. Lisäys 11.7.2018).
Tässä allaolevassa kuvassa on jo tunnettu 3-kinaasi mainittu kuvassa, se kinaasi, jota caffeiini estää ja on hyödyksi antidiabeettisella tavalla.
http://switchgeargenomics.com/wp-content/uploads/2009/08/insulin-receptor-pic.jpg
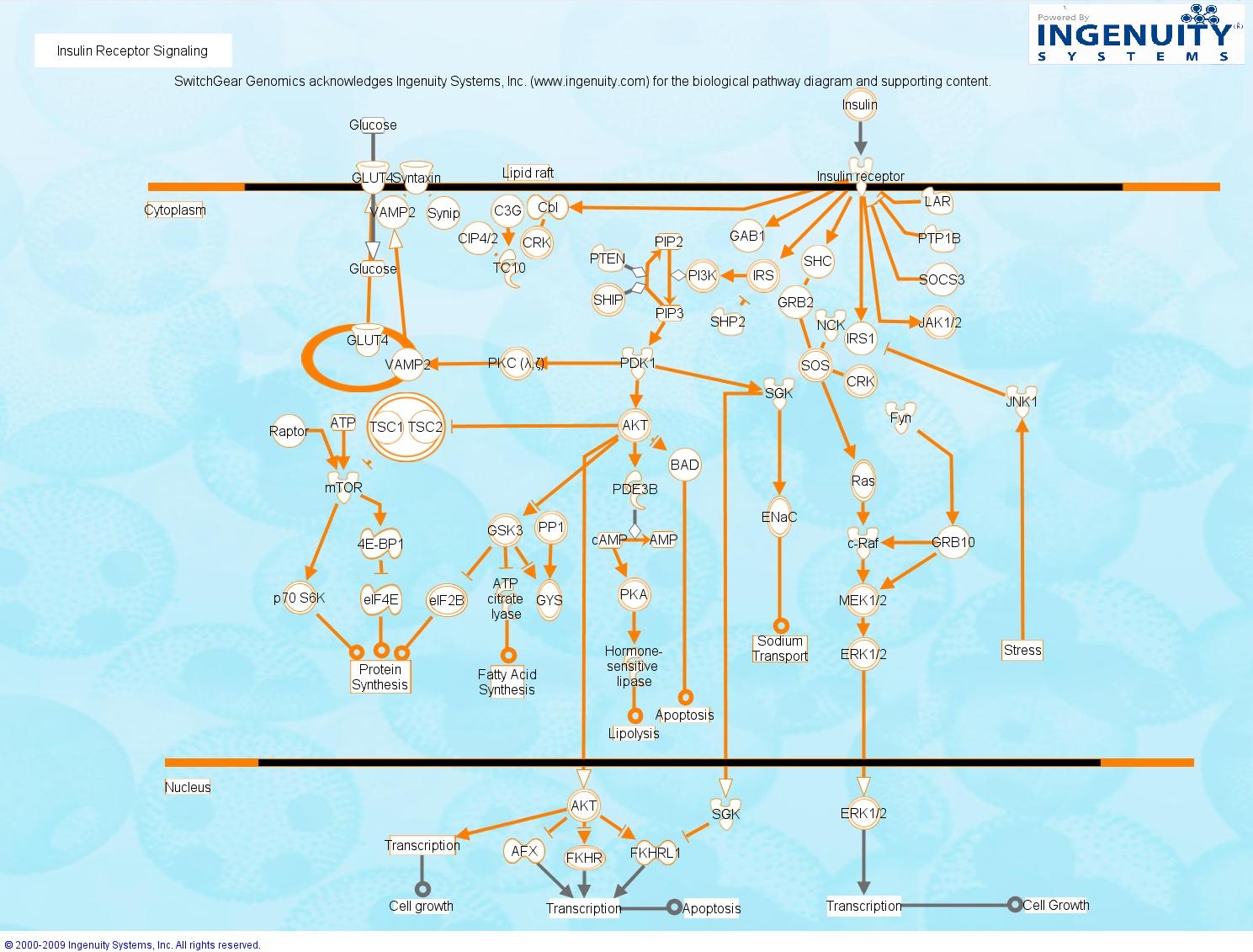{kind=link}
Meneillään oleva tutkimustyö:
Berggren, Per-Olof. Signal-transduction in the pancreatic beta cell.
Karolinska Institutet - Institutionen för molekulär medicin och kirurgi. 2008-11-04
Suomennan tutkijan projektin alustusta.
(1) Työn tarkoituksena on saada ymmärrystä niistä solumekanismeista, jotka säätelevät insuliinin erittymistä beeta-soluista normaalioloissa ja miksi mekanismit eivät toimi diabeteksessa.
- Målsättningen med det aktuella forskningsprojektet är att förstå de cellulära mekanismer som reglerar frisättningen av insulin från våra beta celler under normala betingelser och varför dessa inte fungerar vid diabetes.
(2) Tässä yhteydessä on eräitä tärkeitä asioita selvitettävänä.
Ensinnäkin täytyy ymmärtää
· ne signaalit, joita verensokeri kehkeyttää
· sekä ne eri aineet, joita erittyy muista kuin beetasoluista haiman hormoneita vapauttavasta osasta ja ympäröivistä hermoista
· ja myös ne aineet, jotka pääsevät beetasoluun verisuonisysteemin kautta.
- I detta sammanhang finns det några viktiga saker att klargöra. För det första så måste vi förstå de signaler som genereras av socker i blodet samt av de olika ämnen som frisätts från icke-beta celler i den hormonfrisättande delen av bukspottkörteln och omgivande nerver såväl som av de ämnen som når beta cellen via blodkärlssystemet.
(3) Näiden signaalien täytyy tulla beetasolun integroimiksi, jotta syntyisi ne edellytykset, mitä tarvitaan insuliinin muodostumiseen ja sen vapauttamiseen.
- Dessa signaler måste integreras av beta cellen för att skapa de förutsättningar som behövs för att finjustera såväl bildningen som frisättningen av insulin.
(4) Toiseksi on käsitettävä niitä solumekanismeja, jotka aktivoituvat, kun beetasolu in integroimassa eri signaaleita.
- För det andra så måste vi förstå de cellulära mekanismer som aktiveras då beta cellen integrerar dessa olika signaler.
(5) Kolmanneksi on ymmärrettävä, miten beetasolu toimii elävässä organismissa, kun haimarauhasen hormoneja erittävästa osasta on huolehtimassa sekä verisuonet että hermot.
- För det tredje så måste vi förstå hur beta cellen fungerar i den levande organismen där den hormonfrisättande delen av bukspottkörteln är försörjd av såväl blodkärl som nerver.
(6) Tämän takia tutkijan mielenkiinnon kohteena on seuraavat seikat.
- Därför kommer jag att fokusera mitt intresse på att klargöra
1. Solufunktiolle spesifiset ja tärkeät valkuaisaineet, proteiinit tai jonikanavat, jotka kuljettavat varauksellisia Ca++ atomeita (kalsiumjoneja) beeta-soluun ja miten nämä käyttäytyvät normaaliolosuhteissa ja diabeteksessa.
- Hur specifika och viktiga äggviteämnen för cellulär funktion, proteiner eller jonkanaler, som transporterar laddade kalciumatomer, kalciumjoner, in i beta cellen beter sig under normala betingelser samt vid diabetes.
Hur kalciumjonsignalen beter sig och vad detta betyder för beta cellens funktion och överlevnad.
3. Kuinka varsinainen insuliinia vapauttava koneisto vaikuttuu, jos manipuloidaan joittenkin avaintekijöitten saatavuutta.
- Hur själva det maskineri som reglerar frisättningen av insulin påverkas om man manipulerar tillgängligheten på vissa nyckelspelare.
4. Kuinka muodostuu eräät spesifiset aineet, joita sanotaan inositolipyrofosfaateiksi ja mikä merkitys näillä on beetasolujen funktiolle ja elossapysymiselle.
- Hur specifika ämnen som heter inositol pyrofosfater bildas och vilken betydelse dessa har för beta cellens funktion och överlevnad.
- Hur beta cellen fungerar i den levande organismen.
- För detta ändamål kommer vi att använda en ny teknik som vi nyligen har utvecklat (Nature Medicine on line, publication 7 March 2008) och som gör det möjligt att i detalj studera alla aspekter av beta cells funktion och överlevnad i den levande organismen utan några ingrepp på försöksdjuret och under lång tid. Att på cellulär nivå och i den levande organismen förstå hur den insulinfrisättande beta cellen fungerar är ett måste om man skall kunna utveckla nya mediciner och behandlingsprinciper mot diabetes.
2009-12-11 11:26
(Kommenttini: Tutkija on oikeilla jäljillä. Toivon menestystä!)
Betaolun funktiossakin inositoliaineenvaihdunta on kuin Spanska trappan Roomassa. Kun niitä aikansa nousee, pääsee ylätasanteelle (= pyrofosfaattien integroitu säätely), jossa on Villa Medici ja valtavan suuri paratiisin puu) ja saa hyvät näköalat asiaan- Roomaan... tarkoitan, että kun joka rappu onnistuu jotenkuten, niin että tulee lopulta ylätasanteelle, diabeettista solua ei ole, vaan keho toimii normaalisti, sen diabeettisuus on ylitetty ongelma.
Mutta inositolijärjestelmän jokainen molekyyli ja entsyymi on yksi niistä rapuista, ja jos nyt vielä hokee kerran: Pitäisi tehdä elintarviketaulukot fytiinistä ja inositolista, niin tulevaisuus helpottuisi terapiamenetelmien suhteen sitten aikanaan. Inositoli kilpailee sokeriaineenvaihdunnan kanssa jotenkin ja sen exogeenisesti saadun määrän osuus on tuntematon tekijä.
On tuntematonta miten paljon nykyihminen ja entisajan ihminen käyttää/ käytti fytiiniä ravinnossa ja täten saa/ sai niitä valmiita inositolirenkaita kehoon.(entisajan ihmisillä ei ollut inositoliaineenvaihdunnan rasitteena teollsita glukoosi ja muita vastaavia, jotka kilpailevat inositolin kanssa soluliikenteessä. Lisäys 11.7.2018).
torsdag 10 december 2009
Nobel ja telomeeri
KLINIK OCH VETENSKAP. Kliinistä ja tieteellistä
(1) Flera forskningsmål i sikte … inte bara nya cancerterapier
Nobelpriset om telomerers skyddsfunktion kan ge stor klinisk nytta
Useita tutkimuskohteita on tähtäimessä...ei vain uusia syöpäterapioita.Nobelpalkinto telomeerien suojaavasta vaikutukseta antanee suurta kliinistä hyötyä.
Artikkelin kirjoittajat:
Göran Roos, professor, överläkare; goran.roos( schnabel-a) medbio.umu.se
Pia Osterman, forskningsingenjör; båda institutionen för medicinsk biovetenskap, enheten för patologi, Umeå universitet
YHTEENVETOA
(2) Kun oli keksitty TELOMEERIEN rakenne ja TELOMERAASIN funktio, oli saatu tehtyä perustus sellaiselle tutkimuskentälle, joka viime 25 vuoden aikana on laajentunut räjähdyksenomaisesti, mitä julkaistujen artikkeleitten määrään tulee.
Genom upptäckten av telomerens struktur och telomerasets funktion lades grunden för ett forskningsfält, som under de senaste 25 åren vuxit närmast exponentiellt om man ser till antalet publicerade artiklar.
(3) Ensiksi tutkittiin DNA:n toistuvia sekvenssejä( jaksoja) yksisoluisissa organismeissa ja tästä urautui alkuun hyvin laaja sekä kokeellinen että kliinisluontoinen tutkimus.
Det som startade med studier av repetitiva DNA-sekvenser hos en encellig organism har initierat en mycket bred forskning av såväl experimentell som klinisk natur.
(4) Tämä tutkimus on osoittanut, että TELOMEERIT ja TELOMERAASI ovat funktioltaan konservatiivisia eri organismeissa ja että TELOMEERIT ovat tärkeitä solujen luonnollisessa vanhentumisessa.
Denna forskning har visat att telomerer och telomeras är funktionellt konserverade i olika organismer och att telomerer är viktiga för cellers naturliga åldrande.
(5) TELOMERAASIKOMPLEKSIN geenimutaatiot aiheuttavat sairauksia.
Mutationer i telomeraskomplexets gener är sjukdomsframkallande.
(6) Syöpäsolulle on ominaista kyky pitää yllä tiettyä TELOMEERIPITUUTTA; TELOMERAASIKOMPLEKSI onkin potentielli syöpäterapioitten kohde
En central egenskap för cancerceller är förmågan att upprätthålla en viss telomerlängd, och telomeraskomplexet utgör ett potentiellt mål för nya cancerterapier.
(7) Etukäteen julkaistua. Suunniteltu julkaisu 4/2010.
Förhandspublicering. Planerad publicering i Läkartidningen nr 4/2010
(8) Vuoden 2009 Nobelin palkinto fysiologiassa tai lääketieteessä on jaettu Elizabeth H Blackburnin, Carol W Greiderin ja Jack W Szostakin kesken . Palkinto annetaan siitä löydöstä, että kromosomeja suojaa TELOMEERIT ja ENTSYYMI TELOMERAASI. Tämä löytö on suuri läpilyönti kautta biomedisiinisen alueen ja täten sai ansaitun palkkionsa.
2009 års Nobelpris i fysiologi eller medicin – som tilldelats Elizabeth H Blackburn, Carol W Greider och Jack W Szostak för upptäckten av »hur kromosomerna skyddas av telomerer och enzymet telomeras« – belönar fynd som fått stort genomslag inom skilda biomedicinska forskningsfält.
(9) Palkittu löytö tehtiin 1980-luvulla tutkittaessa hiivasoluja ja Tetrahymena-protozooaorganismia, joka on sovelias kromosomirakenteen ja funktion solubiologisiin analyyseihin , koska sillä on runsas määrä sopivia kromosomeja.
Den belönade upptäckten gjordes på 1980-talet under studier av jästceller och Tetrahymena, en protozo med riklig mängd kromosomer lämpliga för cellbiologiska analyser av kromosomers uppbyggnad och funktion.
(10) Ensimmäisissä julkaisuissa kuvattiin kromosomien päätyosien , TELOMEERIEN, rakennetta ja ne näyttivät olevan koostuneet toistuvista DNA-jaksoista, sekvensseistä; ihmisellä jaksona oli TTAGGG/CCCTAA.
I de första publikationerna [1, 2] beskrevs strukturen av kromosomernas ändpartier, telomererna, vilka visades vara uppbyggda av repetitiva DNA-sekvenser([TTAGGG/CCCTAA]n hos människa).
(11) TELOMEERIEN PITUUS vaihtelee eri kromosomien, eri solutyyppien ja eri lajien kesken. Ihmisellä esiintyvät TELOMEERIT ovat keskimäärin 6 000- 10 000 emästä pitkiä.
Telomerernas längd varierar mellan olika kromosomer, i olika celltyper och bland olika arter och är i medeltal hos människa 6 000–10 000 baser långa.
(12) Metafaasissa olevan solun hybridisoinnilla ja fluorokromilla merkatuilla telomeerikoettimilla on saatu käsitystä TEMOMEERIEN asemasta ja niiden pituuden vaihtelusta eri kromosomien päädyissä. Niiden pääasiallisena funktiona on myöhemmin osoittautunut olevan kromosomin suojaaminen hajoamiselta.
In situ-hybridisering av celler i metafas med användning av en fluorokrommärkt telomerprob illustrerar vackert telomerernas position och att de varierar i längd (här = signalstyrka) mellan olika kromosomändar (Figur 1). Deras huvudsakliga funktion har senare visats vara att stabilisera och skydda kromosomerna från nedbrytning.
(13) Samoihin aikoihin suunnilleen osoitettiin (lähinnä sidekudossolujen) soluviljelmillä, että TELOMEERIT lyhenivät jokaisessa solunjakautumisessa, mikä oli ilmiö, jota teoreettisesti oli aiemminkin jo uumoiltu tapahtuvan.
Ungefär samtidigt visade studier av cellkulturer (främst av bindvävsceller) att telomererna förkortas för varje celldelning, ett fenomen som tidigare på teoretiska grunder förutsatts kunna ske [3].
(14) Normaaleissa ihmissoluissa kromosomit nimittäin menettävät 50-200 emäsparia(bp) TELOMEERIPÄÄDYSTÄ jokaisessa solunjakautumisessa johtuen siitä, että DNA-sekvenssiä ei koskaan voida replikoida ihan päätyyn asti, mikä on ns. päätteen replikaation ongelmaa.
I normala humana celler tappar nämligen kromosomerna 50–200 baspar i slutet av telomeren för varje celldelning, beroende på att DNA-sekvensen inte kan replikeras fullt ut, det s k ändreplikationsproblemet (Figur 2).
(15) On ajateltu niin, että TELOMEERIT on koostu roju-DNA:sta, mikä sinänsä ei koodaa mitään proteiinia, mutta joka voi toimia kuin tikittävä kello solun iän suhteen. Mitä lyhempiä TELOMEERIT ovat , sitä useampia solunjakaumia on solut läpikäyneet.
Telomeren kan anses vara uppbyggd av »skräp-DNA«, som inte kodar för något protein men som kan fungera som en »klocka« för cellens ålder. Ju kortare telomererna är, desto fler celldelningar har cellen genomgått.
(16) On myös voitu osoittaa vahvaa yhteyttä lyhyitten TELOMEERIEN ja sen hetken kesken, jolloin normaalit solut lopettavat jakautumisensa; tämä tarkoittaa sitä, että kriittisellä rajalla ei enää salliudu useampia solunjakautumisia ja solu alkaa vanhentua ja lopulta kuolee.
Man kunde också visa en stark koppling mellan korta telomerer och när normala celler slutar dela sig; dvs vid en kritisk gräns tillåts inte fler delningar, och cellen börjar åldras för att slutligen dö.
(17) TELOMERAASI antaa solulle kykyä ikuistua.
Telomeras ger cellen förmåga till evigt liv
(18) Jotta solut voisivat käydä läpi suuren määrän solunjakautumisia, vaaditaan mekanismia, joka YLLÄPITÄÄ TELOMEERIEN PITUUTTA. Tätä mekanismia ei tunnettu ennen vuotta 1985, jolloin Carol Greider ja Elizabeth Blackburn julkaisivat artikklin, jossa he kuvasivat sellaista entsyymiaktiivisuutta Tetrahymenassa , mikä saattoi pidentää ja luoda uusia TELOMERAASIPÄÄTTEITÄ.
För att celler ska kunna genomgå ett stort antal celldelningar krävs en mekanism som gör att telomerernas längd upprätthålls. Denna mekanism var okänd fram till 1985 då Carol Greider och Elizabeth Blackburn publicerade en artikel [4] där de beskrev att enzymaktivitet i proteinextrakt från Tetrahymena kunde förlänga och skapa nya telomerändar (Figur 2).
(19)
Sittemmin tätä aktiivisuutta kutsuttiin nimellä TELOMERAASI. Se osoittautui olevan käänteinen transkriptaasi, siis sen lisäksi että siinä oli proteiiniosa katalyyttisine aktiivisuuksineen , niin siinä oli mukana myös RNA-komponentti, joka toimi mallina sille nukleotidisekvenssille, mitä kromosomipäätyihin lisätään.
Denna aktivitet kom senare att kallas telomeras och visade sig vara ett omvänt transkriptas, dvs förutom en proteindel med katalytisk aktivitet bär det med sig en RNA-komponent som fungerar som en mall för den nukleotidsekvens som adderas till kromosomändarna (Figur 3).
(20) Blackburn ja Greider olivat ensimmäisiä, jotka osoittivat RNA-komponentin funktion ja merkityksen. Lääketieteen alueella on TELOMERAASIN osoittaminen lisännyt osaltaan tuumoritautien ymmärtämistä, mutta TELOMEERIEN BIOLOGIA on ilmeisesti tärkeä myös muiden tautien kehkeytymisessä.
Blackburn och Greider var även de första att påvisa RNA-komponentens funktion och betydelse [5]. Inom medicinen har påvisandet av telomeras bidragit till ökad förståelse för tumörsjukdomar, men telomerers biologi har visats vara viktig även för uppkomst av andra sjukdomar.
(21) TELOMERAASI on varsinaisesti multiproteiini-RNA-kompleksi, jonka rakenneosaset vaihtelevat eri lajien kesken. Aktiivi kompleksi ihmisessä muodostuu katalyyttisen alayksikön (hTERT) dimeereistä, RNA-komponentista (hTR) ja dyskeriinistä. Dyskeriini on proteiini, joka sitoutuu RNA-molekyyliin.
Telomeras är egentligen ett multiprotein-RNA-komplex, vars beståndsdelar varierar mellan olika arter. Det aktiva komplexet hos människa utgörs av dimerer av den katalytiska subenheten (hTERT), RNA-komponenten (hTR) och dyskerin, ett protein som binder till RNA-molekylen.
(22) Mainittujen rakenneosasten lisäksi on osoittautunut, että TELOMERAASIKOMPLEKSIN kanssa voi käydä interaktioon koko joukko muitakin proteiineja.
Förutom dessa beståndsdelar har en mängd olika proteiner visats kunna interagera med telomeraskomplexet.
(23) TELOMERAASIN tärkeyttä yksisoluiselle "kuolemattomiksi" katsotuille organismille osoittaa se, että TELOMERAASIN MUTAATIO, joka aiheuttaa lyhentyneitä TELOMEEREJA, on niille soluille kuolettava.
Att telomeras är viktigt för encelliga organismer, som kan anses odödliga, bevisas av att om man muterar telomeraset förkortas telomererna, och cellerna dör.
(24) TELOMERAASIAKTIIVISUUTTA voidaan osoittaa oelvan spermioissa, blastokystassa ja useimmissa sikiöaikaisissa kudoksissa varhaisrasakuden aikana ja siten se katoaa.
Telomerasaktivitet kan påvisas i spermier, i blastocyststadiet och i de flesta embryonala vävnader tidigt under graviditeten för att därefter försvinna.
(25) TELOMERAASIA ei tavallisesti esiinny aikuisuudessa normaaleissa kehonsoluissa. Poikkeuksena ovat kantasolut ja epiteelin sekä luuytimen progeniittorisolut sekä veren ja imusolmukkeiden aktivoidut lymfosyytit. Säännönmukaan kyse on sellaisista soluista, jotka jatkuvasti uusiutuvat kautta elämän ,joten niille on eduksi saada uusia TELOMEERISEKVENSSIN lisukkeita. TELOMERAASIAKTIVITEETTI on usein ohimenevää ja solujen erilaistuessa tapahtuu sen vaimennussäätö.
Telomeras uttrycks vanligtvis inte i vuxna normala kroppsceller. Undantag är stamceller och progenitorceller i epitel och benmärg samt aktiverade lymfocyter i blod och lymfkörtlar. Som regel rör det sig om celler i vävnader som ständigt förnyas under livet och där det är en fördel att ibland kunna ge ett nytillskott av telomersekvenser. Telomerasaktiviteten är ofta övergående och nedregleras när cellerna differentierar.
(26) Normaalisoluissa havaittavissa oleva TELOMERAASIAKTIVITEETTI on tavallisesti niin heikkoa, että se ei pysty estämään TELOMEERIPITUUDEN lyhenemistä iän lisääntyessä. Yksi poikkeus on ja se on spermatogeneesi , missä TELOMERAASIN aktiivisuus iän lisääntyessä johtaa tiettyyn pitenemiseen spermioitten TELOMEEREISSÄ. Onhan luonnollisesti mitä tärkein seikka, että se DNA, jonka tehtävänä on ylläpitää tulevia sukupolvia, on niin stabiilia kuin mahdollista ja tähän myötävaikuttaa seprmioitten DNA:n tehokas TELOMEERISUOJA.
Den telomerasaktivitet som kan detekteras i normala celler är vanligtvis inte tillräckligt hög för att förhindra förkortning av telomerlängd med stigande ålder. Ett undantag är spermatogenesen, där telomerasets aktivitet med ökande ålder faktiskt leder till viss förlängning av spermiernas telomerer. Det är naturligtvis av yttersta vikt att det DNA som ska bidra till framtida generationer är så stabilt som möjligt, och till detta bidrar ett fungerande »telomerskydd« för spermie-DNA:t.
(27) Prosessissa on mukana useita proteiineja.
Flertal proteiner med i processen
(28)TELOMEERIPITUUDEN solusäätö ( joka on hyvin monimutkainen), ei ole vielä täysin selvitetty asia. TELOMERAASIN lisäksi on koko joukko proteiineja, jotka sitoutuvat TELOMEEREIHIN ja säätelevät niiden pituutta ja TELOMERAASI-entsyymin pääsyä niihin käsiksi.
Den cellulära regleringen av telomerlängden, som är mycket komplex, är i dagsläget inte helt klarlagd. Förutom telomeras finns det ett flertal proteiner som binder till telomererna och reglerar deras längd och tillgänglighet för telomeras.
(29) Kaikkein tunnetuimpia näistä proteiineista on kuusi, jotka muodsotavat SHELTERIN-KOMPLEKSIN (TRF1, TRF2, Rap1, TIN2, TPP1 ja Pot-1). Shelteriinikompleksin koostumus on tärkeä TELOMEERIEN rakenteelle ja avustaa solua erottamaan toimviat kromosomipäätteet DNA-vaurioista.
Bland de mest studerade är sex proteiner som tillsammans bildar shelterinkomplexet (TRF1, TRF2, Rap1, TIN2, TPP1 och Pot-1) [6]. Sammansättningen av shelterinkomplexet är viktig för strukturen av telomererna och hjälper cellen att skilja funktionella kromosomändar från DNA-skador.
(30) TELOMEEREJÄ tavataan kahta konformaatiota, joka avointa tai T-silmukaksi rullautunutta (TELOMERLOOP) . Silmukkarakenteessa TELOMEERIPÄÄTY on omalla TELOMEERISEKVENSSILLÄ ja se tulee siten suojatuksi.
Telomeren kan ses i två konformationer, antingen som »öppen« eller upprullad i en T-loop (telomerloop). I T-loopsstrukturen har telomeränden invaderat sin egen telomersekvens och blir på så vis skyddad.
(31) Tautia-aiheuttavat TELOMERAASIMUTAATIOT.
Sjukdomsframkallande telomerasmutationer
(32) TELOMERAASIN eri komponenttien mutaatiot ovat omanneet vakavia vaikutuksia moniin solutyyppeihin ja elimiin ja osaltaan vaikuttaneet eri sairauksien esiintymistä. Yhteistä näille taudeille on vaikutus luuytimen soluihin, joilla on näissä potilaissa LYHEMMÄT TELOMEERIT kuin vastaavilla terveillä henkilöillä. MUTAATIOT hTR- ja hTERT-geeneissä esiintyy mm potilailla, joilla on diagnoosina idiopaattinen aplastinen anemia tai myelodysplastinen syndrooma.
Mutationer i telomerasets olika komponenter har visat sig ge allvarliga effekter i många celltyper och organ och kan bidra till uppkomst av skilda sjukdomar. Gemensamt för många av dessa sjukdomar är påverkan på benmärgens celler, vilka hos dessa patienter har kortare telomerer än vad motsvarande friska personer har. Mutationer i hTR- eller hTERT-generna förekommer hos bl a patienter med diagnosen idiopatisk aplastisk anemi eller myelodysplastiskt syndrom [7].
(33) Dyskeratosis congenita on eräs monisysteemisäiraus, johon liittyy ihon pigmentoituminen, kynsimuutokset, suun limakalvon leukoplakia, kyynelkanavien surkastuminen, aplastinen anemia, keuhkofibroosi ja miespotilailla kivessurkastuma. Tässä sairaudessa on joko dyskeriinigeenin(DKC1) tai hTERT- tai hTR-geenin mutaatio. Tavallisin on DKC1 geenin mutaatio (35%)
Dyskeratosis congenita är en multisystemsjukdom associerad med kutan pigmentering, nagelförändringar, leukoplaki i munslemhinnan, atresi av tårkanalen, aplastisk anemi, lungfibros och hos manliga patienter som regel testikelatrofi. Vid denna sjukdom är antingen genen för dyskerin (DKC1) eller hTERT- eller hTR-genen muterad. Vanligast är mutationer i DKC1-genen (35 procent) [8].
(34) DKC1-geenimutaatioita löytyy myös potilailta, joilla on Høyeraal–Hreidarssons syndroma Oireyhtymään kuuluu aplastinen anemia, immuunivaje, mikrokefalia ja kasvunhäiriö.
Mutationer i DKC1-genen hittar man även hos patienter med Høyeraal–Hreidarssons syndrom, en multisystemsjukdom som karakteriseras av aplastisk anemi, immunbrist, mikrocefali och tillväxtstörning [9].
(35) idiopaattinen keuhkofibroosi alkaa aikuisuudessa ja johtaa letaaliin keuhkosidekudoksen muuntumiseen. Taudissa on perinnöllisiä ja sporadisia muotoja. Nykyisin on osoitettu, että hTERT- ja hTR-geenit johtavat telomeerien lyhenemiseen, mikä ajan myötä osaltaan lisää herkkyyttä idiopaattisen keuhkofibroosin kehittymiseen.
Idiopatisk lungfibros debuterar i vuxen ålder och leder till letal bindvävsomvandling av lungparenkymet. Såväl ärftliga som sporadiska former av sjukdomen finns. Nyligen har visats att mutationer i hTERT- eller hTR-generna resulterar i telomerförkortning, vilken med tiden bidrar till ökad känslighet för att utveckla idiopatisk lungfibros [10].
(36) TELOMEERIEN ja syövän välinen yhteys.
Koppling mellan telomerer och cancer
(37) Jos kromosomista puuttuu TELOMEERIT, tämä tilanne vastaa DNA-vauriota, kaksoissäikeen vauriota, jota solu koettaa korjata. Eräs tapa korjata tällaista vauriota on asettaa eri kromosomipäädyt yhteen niin, että muodostuu disentrisiä kromosomeja. Solunjakautumisessa niillä on riskinä murtua, jolloin kromosomaalinen materiaali jakautuu tasaisesti tytärsolujen kesken. Täten voi muodostua kromosomipoikkeamia, mistä teoreettisesti ottaen voi syntyä tuumoreita. Tuumoreilla näyttää myös olevan lyhempiä TELOMEEREJÄ kuin vastaavasa nomaalikudoksessa, minkä selittänee, että tuumorisolut ovat käyneet läpi suuremman määrän solunjakautumisia kuin normaalikudos.
Kromosomer utan telomerer kan liknas vid en DNA-skada, ett dubbelsträngsbrott, som cellen försöker att reparera. Ett sätt kan vara att olika kromosomändar sätts ihop så att dicentriska kromosomer bildas. Vid celldelning riskerar de att brytas sönder, varvid det genetiska materialet fördelas ojämnt mellan dottercellerna. På detta vis kan kromosomala avvikelser uppstå, vilka teoretiskt kan bidra till tumöruppkomst. Det har också visat sig att tumörer har kortare telomerer än motsvarande normalvävnad, vilket kunde förklaras med att tumörcellerna genomgått ett jämförelsevis större antal celldelningar än normalvävnaden.
(38) 1990-luvun alussa tutkittiin munuaissyöpää ja oletettiin, että tuumoreitten läpikäymien solusyklien määrä voitaisiin retrospektiivisesti arvioida analysoimalla niiden TELOMEERIEN PITUUS. Tuloksena oli kuten odotettuakin, että useimmillä tuumoreilla oli normaalia munuaiskudosta lyhemmät TELOMEERIT ja tästä tietomäärästä käsin laskettiin läpikäytyjen solusyklien määrä. Ei ollutkuitenkaan yhteyttä solusyklien määrällä ja tuumorin koolla.Tuumorin koko voinee riippua TELOMERAASIAKTIIVISUUDESTA, jota silloin ei vielä oltu osoitettu tuumoreista , vaikkakin kyllä koeputkessa maligneista soluista.
Vi studerade i början av 1990-talet njurcancer och antog att antalet cellcykler som tumörerna hade genomgått skulle kunna bedömas retrospektivt genom att analysera deras telomerlängd. Resultatet var som väntat att de flesta tumörerna hade kortare telomerer än normal njurvävnad, och utifrån dessa data gjordes beräkningar av antal genomgångna celldelningar [11]. Vi fann dock att det inte fanns någon koppling mellan antal beräknade celldelningar och tumörstorlek och föreslog att det kunde bero på telomerasaktivitet, som man då ännu inte påvisat i tumörer men väl i kulturer av maligna celler in vitro.
(39) Samana vuonna osoitettiin ensimmäistä kertaa AKTIIVIA TELOMERAASIA ihmisen maligniteeteissa.
Samma år påvisades också för första gången aktivt telomeras i humana maligniteter [12, 13].
(49)Aktiviteetin osoittaminen vaati konventionellia molekyylibiologista menetelmää, joka vei aikaa, oli suhteellisen epäherkkä ja vaati paljon soluja. Pian sen jälkeen esitettiin PCR-tekniikka, jossa TELOMERAASITUOTE (= vastikään muodostuneet telomeerisekvenssit) saatettiin tehdä moninkertaisiksi ja täten visualisoida. PCR-metodilla on paljon parempi herkkyys ja niinkin vähän kuin 50- 100 telomeraasipositiivista solua voidaan havaita. Sitten edistyttiin nopeasti TELOMEERIBIOLOGISISSA TUTKIMUKSISSA ja erilaisista malliorganismeista saatu tieto on rikastanut ihmissoluista tehtyjä tutkimuksia hyvin positiivisella tavalla.
För att kunna visa denna aktivitet användes konventionell molekylärbiologisk metodik, som var tidsödande, relativt okänslig och krävde många celler. Snart därefter presenterades en PCR-baserad teknik med vilken telomerasprodukten (nybildade telomersekvenser) kan mångfaldigas och därigenom visualiseras [14]. PCR-metoden har mycket högre känslighet, och så lite som 50–100 telomeraspositiva celler kan detekteras. Därefter startade en snabb utveckling av telomerbiologisk forskning, där data från skilda modellorganismer berikat studier av mänskliga celler på ett mycket positivt sätt.
(50) TELOMERAASILLA on kaksoisrooli tuumorin kehityksessä.
Telomeras spelar dubbelroll i tumörutveckling
(51) Vaikuttaa siltä, että TELOMERAASILLA on tuplaroolit.
Toisaalta sen funktiona on estää tuumoreitten muodostuminen siten, että se säädetään useimmissä solutyypeissä vaimentumaan syntymän jälkeen. Sellaiset solut voivat käydä läpi rajallisen määrän solunjakautumisia ennenkuin TELOMEERIT tulevat KRIITTISEN LYHYIKSI ja solut saavat sitten signaaleita, jotka johtavat kasvunpysähtymään.
Telomeras verkar alltså kunna spela dubbla roller. Å ena sidan är dess funktion att förhindra uppkomst av tumörer genom att det nedregleras i de flesta celltyper efter födseln. Dessa celler kan genomgå ett begränsat antal celldelningar innan telomererna blir kritiskt korta och cellen får signaler som leder till tillväxtstopp.
(52) Toisaalta TELOMERAASI antaa osansa tuumorin syntymiseen, kun se aktivoituessaan stabilisoi/pidentää TELOMEERIEN PITUUTTA, mitkä antaa mahdollisuuksia solunjakautumisiin ja periaatteessa rajattomaan kasvuun.
Å andra sidan bidrar telomeras till tumöruppkomst genom att det vid aktivering stabiliserar/förlänger telomerlängden, vilket möjliggör fler celldelningar och i princip obegränsad tillväxt.
(53) Hanahan et Weinberg ovat identifioineen tämän rajattoman replikatiivisen potentiaalin, kyvyn rajattomaan määrään solunjakautumisia. Tähän kykyyn tarvitaan tietyn TELOMEERIPITUUDEN intaktina säilyminen. Monet tutkimukset ovat osoittaneet, että 80-90%:ssa kaikista maligneista tuumoreista esiintyy AKTIIVIA TELOMERAASIA ja siis kykyä pitää TELOMEERIT riittävinä. Kuten yllä mainittiin tuumoreitten TELOMEERIPITUUDET ovat kyllä lyhempia kuin normaalikudoksissa huolimatta TELOMERAASIN läsnäolosta.
I en ofta citerad översiktsartikel identifierade Hanahan och Weinberg [15] »cancerns kännetecken«, och ett av dessa var »obegränsad replikativ potential«, dvs förmåga till obegränsat antal celldelningar. För denna förmåga krävs att en viss telomerlängd behålls intakt, och i ett stort antal studier har visats att 80–90 procent av alla maligna tumörer har aktivt telomeras och alltså förmåga att bibehålla sina telomerer. Som påpekats ovan är telomerlängden i tumörer ofta kortare än i motsvarande normal vävnad, trots närvaro av telomeras.
(54) Tässä voi pohtia, onko TELOMERAASIAKTIIVISUUS myöhäistapahtuma tuumorin kehittymisessä. Eräs mahdollinen skenaario on, että solut ovat hankkineet mutaatioita, jotka stimuloivat niitä kasvamaan kontrolloimattomasti sekä stimuloivat niitä olemaan reagoimatta normaaleihin jarruttaviin signaaleihin. Ne lyhentävät TELOMEERINSA ja hankkivat lisää geneettisiä muuntumisia, jotka voivat johtaa mm TELOMERAASIN aktivoitumiseen. TELOMERAASI sitten stabilisoi lyhyemmät TELOMEERIPÄÄTTEET ja mahdollistaa rajattomat solunjakautumiset
Man kan här spekulera om huruvida telomerasaktivering är en sen händelse i tumörutvecklingen. Ett tänkbart scenario är att cellerna erhåller mutationer som stimulerar dem att växa okontrollerat och inte reagera på normala bromsande signaler. De förkortar sina telomerer och erhåller fler genetiska förändringar, som kan leda till bl a aktivering av telomeras. Telomeraset stabiliserar härefter den korta telomerlängden och möjliggör obegränsad celldelning.
(55) TELOMERAASINEGATIIVISET tuumorit ovat siis epätavallisia ja usein ne ovat kehittäneet toisen mekanimsin pitääkseen yllä TELOMERAASIPITUUTTA. Tällainen mekanismi rakentuu homologiin rekombinaatioon ja sitä kutsutaan ALT (alternatiivi TELOMEERIEN pidentyminen) . ALT-mekanismia nähdään ennenkaikkea sarkoomassa ja aivotuumoreissa. Ei olla selvillä siitä, miten suuri osa telomeraasinegatiivisista tuumoreista käyttää ALT ja pystyykö molemmat mekanismit( ALT ja TELOMERAASI) toimimaan samaan aikaan tuumoreissa. In vitro( koeputkessa) voidaan kombinoida nämä kaksi järjestelmää.
Telomerasnegativa tumörer är alltså ovanliga, och de har ofta utvecklat en annan mekanism för att upprätthålla telomerlängden. Denna mekanism bygger på homolog rekombination och kallas ALT (alternative lengthening of telomeres). ALT-mekanismen ses framför allt i sarkom och hjärntumörer. Det är inte känt hur stor andel av de telomerasnegativa tumörerna som använder sig av ALT och huruvida bägge mekanismerna (ALT och telomeras) kan samexistera i tumörer. In vitro, i cellinjer, har man kunnat kombinera dessa två system.
(56) Moni tutkimus selvittää ihmistuumorien TELOMERAASIAKTIIVISUUTTA suhteessa taudin kliiniseen kulkuun ja enusteeseen. Tulokset ovat ristiriitaisia useimpien tuumorimuotojen shteen ja varhaiset tiedot yhteyksistä korkean telomeraasiaktiivisuuden ja huonon ennusteen kesken eivät näytä pitävän paikkaansa.
Ett stort antal studier har analyserat telomerasaktivitet i humana tumörer i relation till kliniskt förlopp och prognos. Resultaten är motstridiga för de flesta tumörformer, och tidiga data som fann en koppling mellan höga telomerasnivåer och dålig prognos har inte visat sig hålla.
(57) TELOMERAASIPITUUS on ehkä tärkeämpi seikka kuin TELOMERAASIAKTIIVISUUS.
Telomerernas längd kanske viktigare än telomerasaktivitet
(58) Tuumorisoluissa näyttää olevan tärkeämpi seikka TELOMEERIEN PITUUS eikä niinkään TELOMERAASIN AKTIIVISUUS: Tämä pätee erityisesti hematopoieettisiin maligniteetteihin, joista saadun tiedon mukaan lyhyitä TELOMEEREJÄ vastaa progressiivinen tauti.
En viktigare faktor än telomerasaktivitet verkar den faktiska telomerlängden i tumörcellerna vara. Detta gäller speciellt hematopoetiska maligniteter för vilka ett genomgående fynd är att korta telomerer är associerade med progressiv sjukdom.
(59) Tutkituin tauti tässä suhteessa on KLL, krooninen lymfaattinen leukemia, jossa TELOMERAASIPITUUS( ja KORKEA TELOMERAASIAKTIIVISUUS) on osoittautunut riippumattomaksi prognostiseksi merkitsijäksi. Vastaavankaltainen löytö on tehty multippelista myeloomasta ja myelodysplastisesta oireyhtymästä. Tässä indisoi lyhyet TELOMEERIT pikaisempaa kehitystä kohti akuuttia leukemiaa. Ref. 16 antaa yleiskatsausta telomeeripituudesta biologisena merkitsijänä maligniteetissa.
Den mest studerade sjukdomen i detta avseende är kronisk lymfatisk leukemi där telomerlängd (och hög telomerasaktivitet) visats vara en oberoende prognostisk biomarkör. Liknande fynd har gjorts vid multipelt myelom och myelodysplastiskt syndrom. Här indikerar korta telomerer snabbare utveckling till akut leukemi. I referens 16 ges en översikt av telomerlängd som biomarkör vid malignitet.
(60) TELOMERAASI syöpäterapiassa kohteena. Telomeras som mål för cancerterapi
(61) Koska TELOMERAASI on aktivoituneena useimmissa maligneissa tuumoreissa ja puuttuu useimmista normaaleista soluista, on TELOMERAASI teoreettisesti ottaen kiehtova syöpäterapian kohde. Erilaisia menetelemiä on jo kokeellisissa ja kliinisissa vaiheissa. TELOMERAASIN ESTON pitäisi johtaa TELOMEERIEN LYHENEMISEEN ja lopulta solukuolemaan, mutta tarvittaneen aivan liian monta solunjakaumaa ennenkuin tällainen efekti saavutettaisiin. Tämän takia on ajateltu käyttää TELOMERAASINESTÄJÄÄ kirurgian ja solumyrkkyhoidon jälkeen "käsittelemään" jäljellä jääneitä tuumorisoluja. On päätelty, että normaalit solut, joissa on ohimenevästi aktivoitunutta telomeraasia , eivät vaikuttuisi vakavasti sellaisesta käsittelystä.
Eftersom telomeras är aktiverat i merparten av maligna tumörer och saknas i de flesta normala celler, är telomeras ett teoretiskt attraktivt mål för cancerterapi. Skilda tillvägagångssätt testas nu experimentellt och i kliniska försök [17]. Hämning av telomeras bör leda till telomerförkortning och slutligen cellens död, men det kan behövas (alltför) många celldelningar innan denna effekt uppnås. Därför har en tanke varit att använda telomerashämmare efter kirurgi och cellgiftsbehandling för att »ta hand om« eventuella kvarvarande tumörceller. Det har bedömts att normala celler med övergående aktivering av telomeras inte påverkas allvarligt av sådan behandling.
(62) Voidaan harkita tulevaisia terapeuttisia järjestelmiä, joissa vaikutus on suoraan toksinen niissä soluissa jotka ilmentävät TELOMERAASIA. Kliinisissä kokeissa on nykyhetkellä TELOMERAASIN INHIBIITTOREITA ja myös TERT- immuuniterapiaa, missä ajatuksena on stimuloida immuunivastetta ja "siivota pois" tuumorisoluja, jotka ilmentävät TELOMERAASIA.
Man kan även tänka sig framtida terapeutiska ansatser, där effekten är direkt toxisk i celler som uttrycker telomeras. Kliniska försök pågår i dagsläget med telomerasinhibitorer och även med TERT-immunterapi, där tanken är att stimulera immunförsvaret att »städa bort« tumörceller som uttrycker telomeras.
(63) TELOMEERIEN biologia kytkeytyy vahvasti solujen vanhenemiseen.
Telomerernas biologi starkt kopplad till cellers åldrande
(64) Kute yllä on viitattu soluviljelmissä on vahvaa yhteyttä TELOMEERIEN LYHENEMÄN ja solujakautumisten lukumäärän kesken. Samalla tavalla on vanhemmillä henkilöillä veriroluissaan LYHEMPIÄ TELOMEEREJÄ kuin nuorilla. On myös tavallaista, että naisilla on jonkin verran PITEMPIÄ TELOMEEREJÄ kuin miehillä , mistä on siten tehty yhteyksiä naisten ja miesten elinikään. Näyttää siltä, että naiset menettävät TELOMEERISEKVENSSIÄ kuin miehet jokaisessa hematopoieesin solunjakautumassa.
Som påpekats ovan finns i cellkulturer en stark koppling mellan telomerförkortning och antal celldelningar. På samma sätt ses kortare telomerer i blodceller hos äldre individer än hos yngre personer. Det är också så att kvinnor i sina blodceller har något längre telomerer än män, varför det har varit naturligt att göra en koppling till kvinnors och mäns livslängd. Kvinnor tycks således förlora mindre telomersekvens än män för varje celldelning inom hematopoesen.
(65) Geneettisesti modifioiduilla hiirillä, joilta puuttuu toimiva TELOMERAASI, ei havaita aluksi mitään muutoksia. Tämä johtuu siitä, että hiirillä on aluksi HYVIN PITKÄT TELOMEERIT ja vasta neljännessä sukupolvessa tulee esiin niin lyhyitä TELOMEEREJA, että fenotyyppisiä muutoksia on havaittavissa. Silloin hiirillä on harmaa turkki, huono haavojen paraneminen, limakalvojen surkastumaa maha-suolikanavassa, huonontuneet veriarvot ja steriliteettiä, muutoksia, jotka muistuttavat osittain normaalia vanhenemista.
Genetiskt modifierade möss som saknar fungerande telomeras visar initialt inga förändringar. Detta beror på att möss har mycket långa telomerer, och först efter fyra generationer har telomererna blivit så korta att fenotypiska förändringar uppträder. Mössen visar då gråhårig päls, dålig sårläkning, slemhinneatrofi i mag–tarmkanalen, försämrade blodvärden och sterilitet, förändringar som delvis påminner om naturligt åldrande.
(66) On tauteja, joissa yhtenä oireena on varhainen vanheneminen. Nämä potilaat omaavat lyhempiä TELOMEEREJA kuin vastaavat terveet. Näiden sairauksien taustana vaikuttaa olevan mutaatiot sellaisissa geeneissä, jotka avustavat DNA-vaurion korjaantumista.
Det finns sjukdomar där ett av symtomen är för tidigt åldrande (ataxi–telangiektasisyndromet, Werners syndrom, Fanconis anemi), och dessa patienter uppvisar kortare telomerer än motsvarande friska individer. Bakomliggande orsak för dessa sjukdomar verkar vara mutationer i gener som bidrar till reparation av DNA-skador.
(67) TELOMEERIEN BIOLOGIA on siis vahvasti liittynyt solujen replikatiivisen kykyyn ja solujen ikääntymiseen, mutta ikääntymisprosessi on orgaanitasossa paljon monimutkaisempaa ja siihen vaikuttaa joukko erilaisia tekijöitä.
Telomerernas biologi är således starkt associerad med cellers replikativa förmåga och med cellulärt åldrande, men på organismnivå är åldrandeprocessen mycket mer komplicerad och påverkas av en rad skilda faktorer.
(68) Verisolujen TELOMEERIPITUUS toimii biologisena merkitsijänä.
Blodcellers telomerlängd fungerar som biomarkör
(69) Ihmisen verisoluista on tehty paljon TELOMEERIPITUUTTA koskevia tutkimuksia, samoin telomeeripituuden suhteesta patofysiologisiin ja epidemiologisiin tekijoihin ja eri sairauksiin. Lyhemmät TELOMEERIT potilailla kontrolleihin verrattuna tavataan esim hypertoniassa, diabeteksessa, syövässä, lihavuudessa, verenlipidien epänormaaliuksissa, pitkässä psyykkisessä rasituksessa, inaktiviteetissa ja tupakanpolttajilla. Eri tutkimustulokset ovat toisistaan poikkeavia jonkin verran eikä olla tultu yksimielisyyteen oikeastaan näistä minkään tekijän suhteen.
För människa finns det många studier av telomerlängd i blodceller och dess relation till patofysiologiska och epidemiologiska faktorer samt till skilda sjukdomar. Kortare telomerer hos patienter än i matchade kontrollmaterial har beskrivits vid t ex hypertoni, diabetes, cancer, fetma, abnorma blodlipider, långvarig psykisk belastning, inaktivitet och hos rökare. Skilda publikationer är dock inte helt överensstämmande, och konsensus har knappats nåtts helt avseende någon av dessa faktorer.
(70) Monessa tilassa/faktorissa yhteinen nimittäjä, joka on yhdistettävissä lyhyisiin TELOMEEREIHIN, on oksidatiivinen stressi , mikä on tunnettu tekijä aiheuttamassa lisääntyneitä TEMOMEERIMENETYKSIÄ. Epidemiologiset analyysit ovat vaikeita tehdä, koska varhaisen elämän TELOMEERIPITUUS määräytyy perimän mukaan, lähinnä isän puolelta, ja myöhemmin vaikuttuu elämäntyylistä ja ympäristötekijöistä. Lisäksi eräs komplisoiva tekijä on sekin, että YKSILÖLLINEN TELOMEERIPITUUS osittain määräytyy TELOMEERIEN ALKUPITUUDESTA siten, että pitkät TELOMEERIT omaavalla yksilöllä on tietyssä ajanjaksossa kaikkein huomattavin TELOMEERIEN LYHENTYMINEN
En gemensam nämnare för många tillstånd/faktorer som kopplats till kortare telomerer är oxidativ stress, vilket är en känd faktor som kan bidra till ökade telomerförluster. Analyser av epidemiologisk natur är svåra, eftersom vår telomerlängd tidigt i livet till stor del bestäms av arvet (främst från fadern) för att senare påverkas av livsstil och omgivningsfaktorer. Ytterligare en komplicerande faktor är att den individuella telomerförlusten till en del bestäms av ursprungslängden på telomererna, på så sätt att individer med långa telomerer visat den mest uttalade telomerförkortningen under en viss tidsperiod.
(71) Hämmentävä, mutta mielenkiintoinen löytö on sekin, että veren TELOMEERIPITUUS voi olla prognostinen tekijä syövässä. Prognoosi on huonompi niillä, joilla on pitkät telomeeritperifeerisen veren "buffy-coat-soluissa ( jyväisissä valkosoluissa , imusoluissa ja monosyyteissä). Tämä on havaittu rinta-ja munuaissyöpäpotilailta. Molemmsisa ryhmissä oli telomeeripituus riippumaton prognostinen indikaattori. Nämä tiedot osoittavat, että TELOMEERIPITUUS veressä ei ole staattinen, vaan voi vaikuttua tuumoritaudista. Näiden muutosten taustamekanismit ovat monimutkaisia eikä niitä ole vielä selvitetty.
Ett förvånande men intressant fynd är att telomerlängd i blod kan vara en prognostisk faktor vid cancer. Den patientgrupp som hade långa telomerer i buffy coat-celler (granulocyter, lymfocyter och monocyter) från perifert blod befanns nämligen ha sämre prognos än övriga patienter. Detta resultat har erhållits från patienter med såväl bröst- som njurcancer. I båda patientgrupperna var telomerlängd en signifikant oberoende prognostisk indikator [16]. Dessa data tyder ytterligare på att telomerlängden i blod inte är statisk utan kan påverkas, som i detta fall av tumörsjukdom. Mekanismerna bakom dessa förändringar är komplexa och har ännu inte klarlagts.
Kirjoittajien antamat viitteet:
Referenser
1. Shampay J, Szostak JW, Blackburn EH. DNA sequences of telomeres maintained in yeast. Nature. 1984;310(5973):154-7.
2. Szostak JW, Blackburn EH. Cloning yeast telomeres on linear plasmid vectors. Cell. 1982;29(1):245-55.
3. Harley CB, Futcher AB, Greider CW. Telomeres shorten during ageing of human fibroblasts. Nature. 1990;345(6274):458-60.
4. Greider CW, Blackburn EH. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell. 1985;43:405-13.
5. Greider CW, Blackburn EH. A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis. Nature. 1989;337(6205):331-7.
6. de Lange T. Shelterin: the proteincomplex that shapes and safeguards human telomeres. Genes Dev. 2005; 19(18):2100-10.
7. Yamaguchi H, Baerlocher GM, Lansdorp PM, Chanock SJ, Nunez O, Sloand E, et al. Mutations of the human telomerase RNA gene (TERC) in aplastic anemia and myelodysplastic syndrome. Blood. 2003;102(3):916-8.
8. Mitchell JR, Wood E, Collins K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature. 1999;402(6761):551-5.
9. Knight SW, Heiss NS, Vulliamy TJ, Aalfs CM, McMahon C, Richmond P, et al. Unexplained aplastic anaemia, immunodeficiency, and cerebellar hypoplasia (Hoyeraal-Hreidarsson syndrome) due to mutations in the dyskeratosis congenita gene, DKC1. Br J Haematol. 1999;107(2):335-9.
10. Tsakiri KD, Cronkhite JT, Kuan PJ, Xing C, Raghu G, Weissler JC, et al. Adult-onset pulmonary fibrosis caused by mutations in telomerase. Proc Natl Acad Sci U S A. 2007;104(18):7552-7.
11. Mehle C, Ljungberg B, Roos G. Telomere shortening in renal cell carcinoma. Cancer Res. 1994;54(1):236-41.
12. Counter CM, Hirte HW, Bacchetti S, Harley CB. Telomerase activity in human ovarian carcinoma. Proc Natl Acad Sci U S A. 1994;91(8):2900-4.
13. Nilsson P, Mehle C, Remes K, Roos G. Telomerase activity in vivo in human malignant hematopoietic cells. Oncogene. 1994;9(10):3043-8.
14. Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL, et al. Specific association of human telomerase activity with immortal cells and cancer. Science. 1994;266:2011-5.
15. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100(1):57-70.
16. Svenson U, Roos G. Telomere length as a biological marker in malignancy. Biochim Biophys Acta. 2009;1792(4):317-23.
17. Harley CB. Telomerase and cancer therapeutics. Nat Rev Cancer. 2008;8(3);167-79.
KUVAT
Figur 1. Fluorescent in situ-hybridisering (FISH) av normala celler i metafas och interfas. Signalstyrkan är proportionell mot telomerlängden. FISH-preparation: Sofie Degerman, institutionen för medicinsk biovetenskap, Umeå universitet, och Katarina Nordfjäll, ST-läkare, Östersunds sjukhus.
Figur 2. Normala celler förlorar sina telomerer, och maligna, telomeraspositiva celler behåller sina telomerer för varje celldelning. Utan telomeras förkortas kromosomen varje gång cellen delar sig. Slutligen bryts den ner. Telomeras förlänger telomeren och säkerställer därmed att kromosomen inte blir kortare. Grafik: Annika Röhl/Nobelkommittén för fysiologi eller medicin
http://www.lakartidningen.se/store/images/9/9508/large/Fig2ny.jpg
Figur 3. Principskiss av hur telomeras förlänger telomerer. Angiven DNA-sekvens för telomeren avser Tetrahymena. Enzymet telomeras arbetar på kromosomens ände; det består av ett protein och en RNA-sekvens. RNA-delen är mall när telomer-DNA bildas i änden på kromosomen. Grafik: Annika Röhl/Nobelkommittén för fysiologi eller medicin
http://www.lakartidningen.se/store/images/9/9509/large/Fig3ny.jpg
(1) Flera forskningsmål i sikte … inte bara nya cancerterapier
Nobelpriset om telomerers skyddsfunktion kan ge stor klinisk nytta
Useita tutkimuskohteita on tähtäimessä...ei vain uusia syöpäterapioita.Nobelpalkinto telomeerien suojaavasta vaikutukseta antanee suurta kliinistä hyötyä.
Artikkelin kirjoittajat:
Göran Roos, professor, överläkare; goran.roos( schnabel-a) medbio.umu.se
Pia Osterman, forskningsingenjör; båda institutionen för medicinsk biovetenskap, enheten för patologi, Umeå universitet
YHTEENVETOA
(2) Kun oli keksitty TELOMEERIEN rakenne ja TELOMERAASIN funktio, oli saatu tehtyä perustus sellaiselle tutkimuskentälle, joka viime 25 vuoden aikana on laajentunut räjähdyksenomaisesti, mitä julkaistujen artikkeleitten määrään tulee.
Genom upptäckten av telomerens struktur och telomerasets funktion lades grunden för ett forskningsfält, som under de senaste 25 åren vuxit närmast exponentiellt om man ser till antalet publicerade artiklar.
(3) Ensiksi tutkittiin DNA:n toistuvia sekvenssejä( jaksoja) yksisoluisissa organismeissa ja tästä urautui alkuun hyvin laaja sekä kokeellinen että kliinisluontoinen tutkimus.
Det som startade med studier av repetitiva DNA-sekvenser hos en encellig organism har initierat en mycket bred forskning av såväl experimentell som klinisk natur.
(4) Tämä tutkimus on osoittanut, että TELOMEERIT ja TELOMERAASI ovat funktioltaan konservatiivisia eri organismeissa ja että TELOMEERIT ovat tärkeitä solujen luonnollisessa vanhentumisessa.
Denna forskning har visat att telomerer och telomeras är funktionellt konserverade i olika organismer och att telomerer är viktiga för cellers naturliga åldrande.
(5) TELOMERAASIKOMPLEKSIN geenimutaatiot aiheuttavat sairauksia.
Mutationer i telomeraskomplexets gener är sjukdomsframkallande.
(6) Syöpäsolulle on ominaista kyky pitää yllä tiettyä TELOMEERIPITUUTTA; TELOMERAASIKOMPLEKSI onkin potentielli syöpäterapioitten kohde
En central egenskap för cancerceller är förmågan att upprätthålla en viss telomerlängd, och telomeraskomplexet utgör ett potentiellt mål för nya cancerterapier.
(7) Etukäteen julkaistua. Suunniteltu julkaisu 4/2010.
Förhandspublicering. Planerad publicering i Läkartidningen nr 4/2010
(8) Vuoden 2009 Nobelin palkinto fysiologiassa tai lääketieteessä on jaettu Elizabeth H Blackburnin, Carol W Greiderin ja Jack W Szostakin kesken . Palkinto annetaan siitä löydöstä, että kromosomeja suojaa TELOMEERIT ja ENTSYYMI TELOMERAASI. Tämä löytö on suuri läpilyönti kautta biomedisiinisen alueen ja täten sai ansaitun palkkionsa.
2009 års Nobelpris i fysiologi eller medicin – som tilldelats Elizabeth H Blackburn, Carol W Greider och Jack W Szostak för upptäckten av »hur kromosomerna skyddas av telomerer och enzymet telomeras« – belönar fynd som fått stort genomslag inom skilda biomedicinska forskningsfält.
(9) Palkittu löytö tehtiin 1980-luvulla tutkittaessa hiivasoluja ja Tetrahymena-protozooaorganismia, joka on sovelias kromosomirakenteen ja funktion solubiologisiin analyyseihin , koska sillä on runsas määrä sopivia kromosomeja.
Den belönade upptäckten gjordes på 1980-talet under studier av jästceller och Tetrahymena, en protozo med riklig mängd kromosomer lämpliga för cellbiologiska analyser av kromosomers uppbyggnad och funktion.
(10) Ensimmäisissä julkaisuissa kuvattiin kromosomien päätyosien , TELOMEERIEN, rakennetta ja ne näyttivät olevan koostuneet toistuvista DNA-jaksoista, sekvensseistä; ihmisellä jaksona oli TTAGGG/CCCTAA.
I de första publikationerna [1, 2] beskrevs strukturen av kromosomernas ändpartier, telomererna, vilka visades vara uppbyggda av repetitiva DNA-sekvenser([TTAGGG/CCCTAA]n hos människa).
(11) TELOMEERIEN PITUUS vaihtelee eri kromosomien, eri solutyyppien ja eri lajien kesken. Ihmisellä esiintyvät TELOMEERIT ovat keskimäärin 6 000- 10 000 emästä pitkiä.
Telomerernas längd varierar mellan olika kromosomer, i olika celltyper och bland olika arter och är i medeltal hos människa 6 000–10 000 baser långa.
(12) Metafaasissa olevan solun hybridisoinnilla ja fluorokromilla merkatuilla telomeerikoettimilla on saatu käsitystä TEMOMEERIEN asemasta ja niiden pituuden vaihtelusta eri kromosomien päädyissä. Niiden pääasiallisena funktiona on myöhemmin osoittautunut olevan kromosomin suojaaminen hajoamiselta.
In situ-hybridisering av celler i metafas med användning av en fluorokrommärkt telomerprob illustrerar vackert telomerernas position och att de varierar i längd (här = signalstyrka) mellan olika kromosomändar (Figur 1). Deras huvudsakliga funktion har senare visats vara att stabilisera och skydda kromosomerna från nedbrytning.
(13) Samoihin aikoihin suunnilleen osoitettiin (lähinnä sidekudossolujen) soluviljelmillä, että TELOMEERIT lyhenivät jokaisessa solunjakautumisessa, mikä oli ilmiö, jota teoreettisesti oli aiemminkin jo uumoiltu tapahtuvan.
Ungefär samtidigt visade studier av cellkulturer (främst av bindvävsceller) att telomererna förkortas för varje celldelning, ett fenomen som tidigare på teoretiska grunder förutsatts kunna ske [3].
(14) Normaaleissa ihmissoluissa kromosomit nimittäin menettävät 50-200 emäsparia(bp) TELOMEERIPÄÄDYSTÄ jokaisessa solunjakautumisessa johtuen siitä, että DNA-sekvenssiä ei koskaan voida replikoida ihan päätyyn asti, mikä on ns. päätteen replikaation ongelmaa.
I normala humana celler tappar nämligen kromosomerna 50–200 baspar i slutet av telomeren för varje celldelning, beroende på att DNA-sekvensen inte kan replikeras fullt ut, det s k ändreplikationsproblemet (Figur 2).
(15) On ajateltu niin, että TELOMEERIT on koostu roju-DNA:sta, mikä sinänsä ei koodaa mitään proteiinia, mutta joka voi toimia kuin tikittävä kello solun iän suhteen. Mitä lyhempiä TELOMEERIT ovat , sitä useampia solunjakaumia on solut läpikäyneet.
Telomeren kan anses vara uppbyggd av »skräp-DNA«, som inte kodar för något protein men som kan fungera som en »klocka« för cellens ålder. Ju kortare telomererna är, desto fler celldelningar har cellen genomgått.
(16) On myös voitu osoittaa vahvaa yhteyttä lyhyitten TELOMEERIEN ja sen hetken kesken, jolloin normaalit solut lopettavat jakautumisensa; tämä tarkoittaa sitä, että kriittisellä rajalla ei enää salliudu useampia solunjakautumisia ja solu alkaa vanhentua ja lopulta kuolee.
Man kunde också visa en stark koppling mellan korta telomerer och när normala celler slutar dela sig; dvs vid en kritisk gräns tillåts inte fler delningar, och cellen börjar åldras för att slutligen dö.
(17) TELOMERAASI antaa solulle kykyä ikuistua.
Telomeras ger cellen förmåga till evigt liv
(18) Jotta solut voisivat käydä läpi suuren määrän solunjakautumisia, vaaditaan mekanismia, joka YLLÄPITÄÄ TELOMEERIEN PITUUTTA. Tätä mekanismia ei tunnettu ennen vuotta 1985, jolloin Carol Greider ja Elizabeth Blackburn julkaisivat artikklin, jossa he kuvasivat sellaista entsyymiaktiivisuutta Tetrahymenassa , mikä saattoi pidentää ja luoda uusia TELOMERAASIPÄÄTTEITÄ.
För att celler ska kunna genomgå ett stort antal celldelningar krävs en mekanism som gör att telomerernas längd upprätthålls. Denna mekanism var okänd fram till 1985 då Carol Greider och Elizabeth Blackburn publicerade en artikel [4] där de beskrev att enzymaktivitet i proteinextrakt från Tetrahymena kunde förlänga och skapa nya telomerändar (Figur 2).
(19)
Sittemmin tätä aktiivisuutta kutsuttiin nimellä TELOMERAASI. Se osoittautui olevan käänteinen transkriptaasi, siis sen lisäksi että siinä oli proteiiniosa katalyyttisine aktiivisuuksineen , niin siinä oli mukana myös RNA-komponentti, joka toimi mallina sille nukleotidisekvenssille, mitä kromosomipäätyihin lisätään.
Denna aktivitet kom senare att kallas telomeras och visade sig vara ett omvänt transkriptas, dvs förutom en proteindel med katalytisk aktivitet bär det med sig en RNA-komponent som fungerar som en mall för den nukleotidsekvens som adderas till kromosomändarna (Figur 3).
(20) Blackburn ja Greider olivat ensimmäisiä, jotka osoittivat RNA-komponentin funktion ja merkityksen. Lääketieteen alueella on TELOMERAASIN osoittaminen lisännyt osaltaan tuumoritautien ymmärtämistä, mutta TELOMEERIEN BIOLOGIA on ilmeisesti tärkeä myös muiden tautien kehkeytymisessä.
Blackburn och Greider var även de första att påvisa RNA-komponentens funktion och betydelse [5]. Inom medicinen har påvisandet av telomeras bidragit till ökad förståelse för tumörsjukdomar, men telomerers biologi har visats vara viktig även för uppkomst av andra sjukdomar.
(21) TELOMERAASI on varsinaisesti multiproteiini-RNA-kompleksi, jonka rakenneosaset vaihtelevat eri lajien kesken. Aktiivi kompleksi ihmisessä muodostuu katalyyttisen alayksikön (hTERT) dimeereistä, RNA-komponentista (hTR) ja dyskeriinistä. Dyskeriini on proteiini, joka sitoutuu RNA-molekyyliin.
Telomeras är egentligen ett multiprotein-RNA-komplex, vars beståndsdelar varierar mellan olika arter. Det aktiva komplexet hos människa utgörs av dimerer av den katalytiska subenheten (hTERT), RNA-komponenten (hTR) och dyskerin, ett protein som binder till RNA-molekylen.
(22) Mainittujen rakenneosasten lisäksi on osoittautunut, että TELOMERAASIKOMPLEKSIN kanssa voi käydä interaktioon koko joukko muitakin proteiineja.
Förutom dessa beståndsdelar har en mängd olika proteiner visats kunna interagera med telomeraskomplexet.
(23) TELOMERAASIN tärkeyttä yksisoluiselle "kuolemattomiksi" katsotuille organismille osoittaa se, että TELOMERAASIN MUTAATIO, joka aiheuttaa lyhentyneitä TELOMEEREJA, on niille soluille kuolettava.
Att telomeras är viktigt för encelliga organismer, som kan anses odödliga, bevisas av att om man muterar telomeraset förkortas telomererna, och cellerna dör.
(24) TELOMERAASIAKTIIVISUUTTA voidaan osoittaa oelvan spermioissa, blastokystassa ja useimmissa sikiöaikaisissa kudoksissa varhaisrasakuden aikana ja siten se katoaa.
Telomerasaktivitet kan påvisas i spermier, i blastocyststadiet och i de flesta embryonala vävnader tidigt under graviditeten för att därefter försvinna.
(25) TELOMERAASIA ei tavallisesti esiinny aikuisuudessa normaaleissa kehonsoluissa. Poikkeuksena ovat kantasolut ja epiteelin sekä luuytimen progeniittorisolut sekä veren ja imusolmukkeiden aktivoidut lymfosyytit. Säännönmukaan kyse on sellaisista soluista, jotka jatkuvasti uusiutuvat kautta elämän ,joten niille on eduksi saada uusia TELOMEERISEKVENSSIN lisukkeita. TELOMERAASIAKTIVITEETTI on usein ohimenevää ja solujen erilaistuessa tapahtuu sen vaimennussäätö.
Telomeras uttrycks vanligtvis inte i vuxna normala kroppsceller. Undantag är stamceller och progenitorceller i epitel och benmärg samt aktiverade lymfocyter i blod och lymfkörtlar. Som regel rör det sig om celler i vävnader som ständigt förnyas under livet och där det är en fördel att ibland kunna ge ett nytillskott av telomersekvenser. Telomerasaktiviteten är ofta övergående och nedregleras när cellerna differentierar.
(26) Normaalisoluissa havaittavissa oleva TELOMERAASIAKTIVITEETTI on tavallisesti niin heikkoa, että se ei pysty estämään TELOMEERIPITUUDEN lyhenemistä iän lisääntyessä. Yksi poikkeus on ja se on spermatogeneesi , missä TELOMERAASIN aktiivisuus iän lisääntyessä johtaa tiettyyn pitenemiseen spermioitten TELOMEEREISSÄ. Onhan luonnollisesti mitä tärkein seikka, että se DNA, jonka tehtävänä on ylläpitää tulevia sukupolvia, on niin stabiilia kuin mahdollista ja tähän myötävaikuttaa seprmioitten DNA:n tehokas TELOMEERISUOJA.
Den telomerasaktivitet som kan detekteras i normala celler är vanligtvis inte tillräckligt hög för att förhindra förkortning av telomerlängd med stigande ålder. Ett undantag är spermatogenesen, där telomerasets aktivitet med ökande ålder faktiskt leder till viss förlängning av spermiernas telomerer. Det är naturligtvis av yttersta vikt att det DNA som ska bidra till framtida generationer är så stabilt som möjligt, och till detta bidrar ett fungerande »telomerskydd« för spermie-DNA:t.
(27) Prosessissa on mukana useita proteiineja.
Flertal proteiner med i processen
(28)TELOMEERIPITUUDEN solusäätö ( joka on hyvin monimutkainen), ei ole vielä täysin selvitetty asia. TELOMERAASIN lisäksi on koko joukko proteiineja, jotka sitoutuvat TELOMEEREIHIN ja säätelevät niiden pituutta ja TELOMERAASI-entsyymin pääsyä niihin käsiksi.
Den cellulära regleringen av telomerlängden, som är mycket komplex, är i dagsläget inte helt klarlagd. Förutom telomeras finns det ett flertal proteiner som binder till telomererna och reglerar deras längd och tillgänglighet för telomeras.
(29) Kaikkein tunnetuimpia näistä proteiineista on kuusi, jotka muodsotavat SHELTERIN-KOMPLEKSIN (TRF1, TRF2, Rap1, TIN2, TPP1 ja Pot-1). Shelteriinikompleksin koostumus on tärkeä TELOMEERIEN rakenteelle ja avustaa solua erottamaan toimviat kromosomipäätteet DNA-vaurioista.
Bland de mest studerade är sex proteiner som tillsammans bildar shelterinkomplexet (TRF1, TRF2, Rap1, TIN2, TPP1 och Pot-1) [6]. Sammansättningen av shelterinkomplexet är viktig för strukturen av telomererna och hjälper cellen att skilja funktionella kromosomändar från DNA-skador.
(30) TELOMEEREJÄ tavataan kahta konformaatiota, joka avointa tai T-silmukaksi rullautunutta (TELOMERLOOP) . Silmukkarakenteessa TELOMEERIPÄÄTY on omalla TELOMEERISEKVENSSILLÄ ja se tulee siten suojatuksi.
Telomeren kan ses i två konformationer, antingen som »öppen« eller upprullad i en T-loop (telomerloop). I T-loopsstrukturen har telomeränden invaderat sin egen telomersekvens och blir på så vis skyddad.
(31) Tautia-aiheuttavat TELOMERAASIMUTAATIOT.
Sjukdomsframkallande telomerasmutationer
(32) TELOMERAASIN eri komponenttien mutaatiot ovat omanneet vakavia vaikutuksia moniin solutyyppeihin ja elimiin ja osaltaan vaikuttaneet eri sairauksien esiintymistä. Yhteistä näille taudeille on vaikutus luuytimen soluihin, joilla on näissä potilaissa LYHEMMÄT TELOMEERIT kuin vastaavilla terveillä henkilöillä. MUTAATIOT hTR- ja hTERT-geeneissä esiintyy mm potilailla, joilla on diagnoosina idiopaattinen aplastinen anemia tai myelodysplastinen syndrooma.
Mutationer i telomerasets olika komponenter har visat sig ge allvarliga effekter i många celltyper och organ och kan bidra till uppkomst av skilda sjukdomar. Gemensamt för många av dessa sjukdomar är påverkan på benmärgens celler, vilka hos dessa patienter har kortare telomerer än vad motsvarande friska personer har. Mutationer i hTR- eller hTERT-generna förekommer hos bl a patienter med diagnosen idiopatisk aplastisk anemi eller myelodysplastiskt syndrom [7].
(33) Dyskeratosis congenita on eräs monisysteemisäiraus, johon liittyy ihon pigmentoituminen, kynsimuutokset, suun limakalvon leukoplakia, kyynelkanavien surkastuminen, aplastinen anemia, keuhkofibroosi ja miespotilailla kivessurkastuma. Tässä sairaudessa on joko dyskeriinigeenin(DKC1) tai hTERT- tai hTR-geenin mutaatio. Tavallisin on DKC1 geenin mutaatio (35%)
Dyskeratosis congenita är en multisystemsjukdom associerad med kutan pigmentering, nagelförändringar, leukoplaki i munslemhinnan, atresi av tårkanalen, aplastisk anemi, lungfibros och hos manliga patienter som regel testikelatrofi. Vid denna sjukdom är antingen genen för dyskerin (DKC1) eller hTERT- eller hTR-genen muterad. Vanligast är mutationer i DKC1-genen (35 procent) [8].
(34) DKC1-geenimutaatioita löytyy myös potilailta, joilla on Høyeraal–Hreidarssons syndroma Oireyhtymään kuuluu aplastinen anemia, immuunivaje, mikrokefalia ja kasvunhäiriö.
Mutationer i DKC1-genen hittar man även hos patienter med Høyeraal–Hreidarssons syndrom, en multisystemsjukdom som karakteriseras av aplastisk anemi, immunbrist, mikrocefali och tillväxtstörning [9].
(35) idiopaattinen keuhkofibroosi alkaa aikuisuudessa ja johtaa letaaliin keuhkosidekudoksen muuntumiseen. Taudissa on perinnöllisiä ja sporadisia muotoja. Nykyisin on osoitettu, että hTERT- ja hTR-geenit johtavat telomeerien lyhenemiseen, mikä ajan myötä osaltaan lisää herkkyyttä idiopaattisen keuhkofibroosin kehittymiseen.
Idiopatisk lungfibros debuterar i vuxen ålder och leder till letal bindvävsomvandling av lungparenkymet. Såväl ärftliga som sporadiska former av sjukdomen finns. Nyligen har visats att mutationer i hTERT- eller hTR-generna resulterar i telomerförkortning, vilken med tiden bidrar till ökad känslighet för att utveckla idiopatisk lungfibros [10].
(36) TELOMEERIEN ja syövän välinen yhteys.
Koppling mellan telomerer och cancer
(37) Jos kromosomista puuttuu TELOMEERIT, tämä tilanne vastaa DNA-vauriota, kaksoissäikeen vauriota, jota solu koettaa korjata. Eräs tapa korjata tällaista vauriota on asettaa eri kromosomipäädyt yhteen niin, että muodostuu disentrisiä kromosomeja. Solunjakautumisessa niillä on riskinä murtua, jolloin kromosomaalinen materiaali jakautuu tasaisesti tytärsolujen kesken. Täten voi muodostua kromosomipoikkeamia, mistä teoreettisesti ottaen voi syntyä tuumoreita. Tuumoreilla näyttää myös olevan lyhempiä TELOMEEREJÄ kuin vastaavasa nomaalikudoksessa, minkä selittänee, että tuumorisolut ovat käyneet läpi suuremman määrän solunjakautumisia kuin normaalikudos.
Kromosomer utan telomerer kan liknas vid en DNA-skada, ett dubbelsträngsbrott, som cellen försöker att reparera. Ett sätt kan vara att olika kromosomändar sätts ihop så att dicentriska kromosomer bildas. Vid celldelning riskerar de att brytas sönder, varvid det genetiska materialet fördelas ojämnt mellan dottercellerna. På detta vis kan kromosomala avvikelser uppstå, vilka teoretiskt kan bidra till tumöruppkomst. Det har också visat sig att tumörer har kortare telomerer än motsvarande normalvävnad, vilket kunde förklaras med att tumörcellerna genomgått ett jämförelsevis större antal celldelningar än normalvävnaden.
(38) 1990-luvun alussa tutkittiin munuaissyöpää ja oletettiin, että tuumoreitten läpikäymien solusyklien määrä voitaisiin retrospektiivisesti arvioida analysoimalla niiden TELOMEERIEN PITUUS. Tuloksena oli kuten odotettuakin, että useimmillä tuumoreilla oli normaalia munuaiskudosta lyhemmät TELOMEERIT ja tästä tietomäärästä käsin laskettiin läpikäytyjen solusyklien määrä. Ei ollutkuitenkaan yhteyttä solusyklien määrällä ja tuumorin koolla.Tuumorin koko voinee riippua TELOMERAASIAKTIIVISUUDESTA, jota silloin ei vielä oltu osoitettu tuumoreista , vaikkakin kyllä koeputkessa maligneista soluista.
Vi studerade i början av 1990-talet njurcancer och antog att antalet cellcykler som tumörerna hade genomgått skulle kunna bedömas retrospektivt genom att analysera deras telomerlängd. Resultatet var som väntat att de flesta tumörerna hade kortare telomerer än normal njurvävnad, och utifrån dessa data gjordes beräkningar av antal genomgångna celldelningar [11]. Vi fann dock att det inte fanns någon koppling mellan antal beräknade celldelningar och tumörstorlek och föreslog att det kunde bero på telomerasaktivitet, som man då ännu inte påvisat i tumörer men väl i kulturer av maligna celler in vitro.
(39) Samana vuonna osoitettiin ensimmäistä kertaa AKTIIVIA TELOMERAASIA ihmisen maligniteeteissa.
Samma år påvisades också för första gången aktivt telomeras i humana maligniteter [12, 13].
(49)Aktiviteetin osoittaminen vaati konventionellia molekyylibiologista menetelmää, joka vei aikaa, oli suhteellisen epäherkkä ja vaati paljon soluja. Pian sen jälkeen esitettiin PCR-tekniikka, jossa TELOMERAASITUOTE (= vastikään muodostuneet telomeerisekvenssit) saatettiin tehdä moninkertaisiksi ja täten visualisoida. PCR-metodilla on paljon parempi herkkyys ja niinkin vähän kuin 50- 100 telomeraasipositiivista solua voidaan havaita. Sitten edistyttiin nopeasti TELOMEERIBIOLOGISISSA TUTKIMUKSISSA ja erilaisista malliorganismeista saatu tieto on rikastanut ihmissoluista tehtyjä tutkimuksia hyvin positiivisella tavalla.
För att kunna visa denna aktivitet användes konventionell molekylärbiologisk metodik, som var tidsödande, relativt okänslig och krävde många celler. Snart därefter presenterades en PCR-baserad teknik med vilken telomerasprodukten (nybildade telomersekvenser) kan mångfaldigas och därigenom visualiseras [14]. PCR-metoden har mycket högre känslighet, och så lite som 50–100 telomeraspositiva celler kan detekteras. Därefter startade en snabb utveckling av telomerbiologisk forskning, där data från skilda modellorganismer berikat studier av mänskliga celler på ett mycket positivt sätt.
(50) TELOMERAASILLA on kaksoisrooli tuumorin kehityksessä.
Telomeras spelar dubbelroll i tumörutveckling
(51) Vaikuttaa siltä, että TELOMERAASILLA on tuplaroolit.
Toisaalta sen funktiona on estää tuumoreitten muodostuminen siten, että se säädetään useimmissä solutyypeissä vaimentumaan syntymän jälkeen. Sellaiset solut voivat käydä läpi rajallisen määrän solunjakautumisia ennenkuin TELOMEERIT tulevat KRIITTISEN LYHYIKSI ja solut saavat sitten signaaleita, jotka johtavat kasvunpysähtymään.
Telomeras verkar alltså kunna spela dubbla roller. Å ena sidan är dess funktion att förhindra uppkomst av tumörer genom att det nedregleras i de flesta celltyper efter födseln. Dessa celler kan genomgå ett begränsat antal celldelningar innan telomererna blir kritiskt korta och cellen får signaler som leder till tillväxtstopp.
(52) Toisaalta TELOMERAASI antaa osansa tuumorin syntymiseen, kun se aktivoituessaan stabilisoi/pidentää TELOMEERIEN PITUUTTA, mitkä antaa mahdollisuuksia solunjakautumisiin ja periaatteessa rajattomaan kasvuun.
Å andra sidan bidrar telomeras till tumöruppkomst genom att det vid aktivering stabiliserar/förlänger telomerlängden, vilket möjliggör fler celldelningar och i princip obegränsad tillväxt.
(53) Hanahan et Weinberg ovat identifioineen tämän rajattoman replikatiivisen potentiaalin, kyvyn rajattomaan määrään solunjakautumisia. Tähän kykyyn tarvitaan tietyn TELOMEERIPITUUDEN intaktina säilyminen. Monet tutkimukset ovat osoittaneet, että 80-90%:ssa kaikista maligneista tuumoreista esiintyy AKTIIVIA TELOMERAASIA ja siis kykyä pitää TELOMEERIT riittävinä. Kuten yllä mainittiin tuumoreitten TELOMEERIPITUUDET ovat kyllä lyhempia kuin normaalikudoksissa huolimatta TELOMERAASIN läsnäolosta.
I en ofta citerad översiktsartikel identifierade Hanahan och Weinberg [15] »cancerns kännetecken«, och ett av dessa var »obegränsad replikativ potential«, dvs förmåga till obegränsat antal celldelningar. För denna förmåga krävs att en viss telomerlängd behålls intakt, och i ett stort antal studier har visats att 80–90 procent av alla maligna tumörer har aktivt telomeras och alltså förmåga att bibehålla sina telomerer. Som påpekats ovan är telomerlängden i tumörer ofta kortare än i motsvarande normal vävnad, trots närvaro av telomeras.
(54) Tässä voi pohtia, onko TELOMERAASIAKTIIVISUUS myöhäistapahtuma tuumorin kehittymisessä. Eräs mahdollinen skenaario on, että solut ovat hankkineet mutaatioita, jotka stimuloivat niitä kasvamaan kontrolloimattomasti sekä stimuloivat niitä olemaan reagoimatta normaaleihin jarruttaviin signaaleihin. Ne lyhentävät TELOMEERINSA ja hankkivat lisää geneettisiä muuntumisia, jotka voivat johtaa mm TELOMERAASIN aktivoitumiseen. TELOMERAASI sitten stabilisoi lyhyemmät TELOMEERIPÄÄTTEET ja mahdollistaa rajattomat solunjakautumiset
Man kan här spekulera om huruvida telomerasaktivering är en sen händelse i tumörutvecklingen. Ett tänkbart scenario är att cellerna erhåller mutationer som stimulerar dem att växa okontrollerat och inte reagera på normala bromsande signaler. De förkortar sina telomerer och erhåller fler genetiska förändringar, som kan leda till bl a aktivering av telomeras. Telomeraset stabiliserar härefter den korta telomerlängden och möjliggör obegränsad celldelning.
(55) TELOMERAASINEGATIIVISET tuumorit ovat siis epätavallisia ja usein ne ovat kehittäneet toisen mekanimsin pitääkseen yllä TELOMERAASIPITUUTTA. Tällainen mekanismi rakentuu homologiin rekombinaatioon ja sitä kutsutaan ALT (alternatiivi TELOMEERIEN pidentyminen) . ALT-mekanismia nähdään ennenkaikkea sarkoomassa ja aivotuumoreissa. Ei olla selvillä siitä, miten suuri osa telomeraasinegatiivisista tuumoreista käyttää ALT ja pystyykö molemmat mekanismit( ALT ja TELOMERAASI) toimimaan samaan aikaan tuumoreissa. In vitro( koeputkessa) voidaan kombinoida nämä kaksi järjestelmää.
Telomerasnegativa tumörer är alltså ovanliga, och de har ofta utvecklat en annan mekanism för att upprätthålla telomerlängden. Denna mekanism bygger på homolog rekombination och kallas ALT (alternative lengthening of telomeres). ALT-mekanismen ses framför allt i sarkom och hjärntumörer. Det är inte känt hur stor andel av de telomerasnegativa tumörerna som använder sig av ALT och huruvida bägge mekanismerna (ALT och telomeras) kan samexistera i tumörer. In vitro, i cellinjer, har man kunnat kombinera dessa två system.
(56) Moni tutkimus selvittää ihmistuumorien TELOMERAASIAKTIIVISUUTTA suhteessa taudin kliiniseen kulkuun ja enusteeseen. Tulokset ovat ristiriitaisia useimpien tuumorimuotojen shteen ja varhaiset tiedot yhteyksistä korkean telomeraasiaktiivisuuden ja huonon ennusteen kesken eivät näytä pitävän paikkaansa.
Ett stort antal studier har analyserat telomerasaktivitet i humana tumörer i relation till kliniskt förlopp och prognos. Resultaten är motstridiga för de flesta tumörformer, och tidiga data som fann en koppling mellan höga telomerasnivåer och dålig prognos har inte visat sig hålla.
(57) TELOMERAASIPITUUS on ehkä tärkeämpi seikka kuin TELOMERAASIAKTIIVISUUS.
Telomerernas längd kanske viktigare än telomerasaktivitet
(58) Tuumorisoluissa näyttää olevan tärkeämpi seikka TELOMEERIEN PITUUS eikä niinkään TELOMERAASIN AKTIIVISUUS: Tämä pätee erityisesti hematopoieettisiin maligniteetteihin, joista saadun tiedon mukaan lyhyitä TELOMEEREJÄ vastaa progressiivinen tauti.
En viktigare faktor än telomerasaktivitet verkar den faktiska telomerlängden i tumörcellerna vara. Detta gäller speciellt hematopoetiska maligniteter för vilka ett genomgående fynd är att korta telomerer är associerade med progressiv sjukdom.
(59) Tutkituin tauti tässä suhteessa on KLL, krooninen lymfaattinen leukemia, jossa TELOMERAASIPITUUS( ja KORKEA TELOMERAASIAKTIIVISUUS) on osoittautunut riippumattomaksi prognostiseksi merkitsijäksi. Vastaavankaltainen löytö on tehty multippelista myeloomasta ja myelodysplastisesta oireyhtymästä. Tässä indisoi lyhyet TELOMEERIT pikaisempaa kehitystä kohti akuuttia leukemiaa. Ref. 16 antaa yleiskatsausta telomeeripituudesta biologisena merkitsijänä maligniteetissa.
Den mest studerade sjukdomen i detta avseende är kronisk lymfatisk leukemi där telomerlängd (och hög telomerasaktivitet) visats vara en oberoende prognostisk biomarkör. Liknande fynd har gjorts vid multipelt myelom och myelodysplastiskt syndrom. Här indikerar korta telomerer snabbare utveckling till akut leukemi. I referens 16 ges en översikt av telomerlängd som biomarkör vid malignitet.
(60) TELOMERAASI syöpäterapiassa kohteena. Telomeras som mål för cancerterapi
(61) Koska TELOMERAASI on aktivoituneena useimmissa maligneissa tuumoreissa ja puuttuu useimmista normaaleista soluista, on TELOMERAASI teoreettisesti ottaen kiehtova syöpäterapian kohde. Erilaisia menetelemiä on jo kokeellisissa ja kliinisissa vaiheissa. TELOMERAASIN ESTON pitäisi johtaa TELOMEERIEN LYHENEMISEEN ja lopulta solukuolemaan, mutta tarvittaneen aivan liian monta solunjakaumaa ennenkuin tällainen efekti saavutettaisiin. Tämän takia on ajateltu käyttää TELOMERAASINESTÄJÄÄ kirurgian ja solumyrkkyhoidon jälkeen "käsittelemään" jäljellä jääneitä tuumorisoluja. On päätelty, että normaalit solut, joissa on ohimenevästi aktivoitunutta telomeraasia , eivät vaikuttuisi vakavasti sellaisesta käsittelystä.
Eftersom telomeras är aktiverat i merparten av maligna tumörer och saknas i de flesta normala celler, är telomeras ett teoretiskt attraktivt mål för cancerterapi. Skilda tillvägagångssätt testas nu experimentellt och i kliniska försök [17]. Hämning av telomeras bör leda till telomerförkortning och slutligen cellens död, men det kan behövas (alltför) många celldelningar innan denna effekt uppnås. Därför har en tanke varit att använda telomerashämmare efter kirurgi och cellgiftsbehandling för att »ta hand om« eventuella kvarvarande tumörceller. Det har bedömts att normala celler med övergående aktivering av telomeras inte påverkas allvarligt av sådan behandling.
(62) Voidaan harkita tulevaisia terapeuttisia järjestelmiä, joissa vaikutus on suoraan toksinen niissä soluissa jotka ilmentävät TELOMERAASIA. Kliinisissä kokeissa on nykyhetkellä TELOMERAASIN INHIBIITTOREITA ja myös TERT- immuuniterapiaa, missä ajatuksena on stimuloida immuunivastetta ja "siivota pois" tuumorisoluja, jotka ilmentävät TELOMERAASIA.
Man kan även tänka sig framtida terapeutiska ansatser, där effekten är direkt toxisk i celler som uttrycker telomeras. Kliniska försök pågår i dagsläget med telomerasinhibitorer och även med TERT-immunterapi, där tanken är att stimulera immunförsvaret att »städa bort« tumörceller som uttrycker telomeras.
(63) TELOMEERIEN biologia kytkeytyy vahvasti solujen vanhenemiseen.
Telomerernas biologi starkt kopplad till cellers åldrande
(64) Kute yllä on viitattu soluviljelmissä on vahvaa yhteyttä TELOMEERIEN LYHENEMÄN ja solujakautumisten lukumäärän kesken. Samalla tavalla on vanhemmillä henkilöillä veriroluissaan LYHEMPIÄ TELOMEEREJÄ kuin nuorilla. On myös tavallaista, että naisilla on jonkin verran PITEMPIÄ TELOMEEREJÄ kuin miehillä , mistä on siten tehty yhteyksiä naisten ja miesten elinikään. Näyttää siltä, että naiset menettävät TELOMEERISEKVENSSIÄ kuin miehet jokaisessa hematopoieesin solunjakautumassa.
Som påpekats ovan finns i cellkulturer en stark koppling mellan telomerförkortning och antal celldelningar. På samma sätt ses kortare telomerer i blodceller hos äldre individer än hos yngre personer. Det är också så att kvinnor i sina blodceller har något längre telomerer än män, varför det har varit naturligt att göra en koppling till kvinnors och mäns livslängd. Kvinnor tycks således förlora mindre telomersekvens än män för varje celldelning inom hematopoesen.
(65) Geneettisesti modifioiduilla hiirillä, joilta puuttuu toimiva TELOMERAASI, ei havaita aluksi mitään muutoksia. Tämä johtuu siitä, että hiirillä on aluksi HYVIN PITKÄT TELOMEERIT ja vasta neljännessä sukupolvessa tulee esiin niin lyhyitä TELOMEEREJA, että fenotyyppisiä muutoksia on havaittavissa. Silloin hiirillä on harmaa turkki, huono haavojen paraneminen, limakalvojen surkastumaa maha-suolikanavassa, huonontuneet veriarvot ja steriliteettiä, muutoksia, jotka muistuttavat osittain normaalia vanhenemista.
Genetiskt modifierade möss som saknar fungerande telomeras visar initialt inga förändringar. Detta beror på att möss har mycket långa telomerer, och först efter fyra generationer har telomererna blivit så korta att fenotypiska förändringar uppträder. Mössen visar då gråhårig päls, dålig sårläkning, slemhinneatrofi i mag–tarmkanalen, försämrade blodvärden och sterilitet, förändringar som delvis påminner om naturligt åldrande.
(66) On tauteja, joissa yhtenä oireena on varhainen vanheneminen. Nämä potilaat omaavat lyhempiä TELOMEEREJA kuin vastaavat terveet. Näiden sairauksien taustana vaikuttaa olevan mutaatiot sellaisissa geeneissä, jotka avustavat DNA-vaurion korjaantumista.
Det finns sjukdomar där ett av symtomen är för tidigt åldrande (ataxi–telangiektasisyndromet, Werners syndrom, Fanconis anemi), och dessa patienter uppvisar kortare telomerer än motsvarande friska individer. Bakomliggande orsak för dessa sjukdomar verkar vara mutationer i gener som bidrar till reparation av DNA-skador.
(67) TELOMEERIEN BIOLOGIA on siis vahvasti liittynyt solujen replikatiivisen kykyyn ja solujen ikääntymiseen, mutta ikääntymisprosessi on orgaanitasossa paljon monimutkaisempaa ja siihen vaikuttaa joukko erilaisia tekijöitä.
Telomerernas biologi är således starkt associerad med cellers replikativa förmåga och med cellulärt åldrande, men på organismnivå är åldrandeprocessen mycket mer komplicerad och påverkas av en rad skilda faktorer.
(68) Verisolujen TELOMEERIPITUUS toimii biologisena merkitsijänä.
Blodcellers telomerlängd fungerar som biomarkör
(69) Ihmisen verisoluista on tehty paljon TELOMEERIPITUUTTA koskevia tutkimuksia, samoin telomeeripituuden suhteesta patofysiologisiin ja epidemiologisiin tekijoihin ja eri sairauksiin. Lyhemmät TELOMEERIT potilailla kontrolleihin verrattuna tavataan esim hypertoniassa, diabeteksessa, syövässä, lihavuudessa, verenlipidien epänormaaliuksissa, pitkässä psyykkisessä rasituksessa, inaktiviteetissa ja tupakanpolttajilla. Eri tutkimustulokset ovat toisistaan poikkeavia jonkin verran eikä olla tultu yksimielisyyteen oikeastaan näistä minkään tekijän suhteen.
För människa finns det många studier av telomerlängd i blodceller och dess relation till patofysiologiska och epidemiologiska faktorer samt till skilda sjukdomar. Kortare telomerer hos patienter än i matchade kontrollmaterial har beskrivits vid t ex hypertoni, diabetes, cancer, fetma, abnorma blodlipider, långvarig psykisk belastning, inaktivitet och hos rökare. Skilda publikationer är dock inte helt överensstämmande, och konsensus har knappats nåtts helt avseende någon av dessa faktorer.
(70) Monessa tilassa/faktorissa yhteinen nimittäjä, joka on yhdistettävissä lyhyisiin TELOMEEREIHIN, on oksidatiivinen stressi , mikä on tunnettu tekijä aiheuttamassa lisääntyneitä TEMOMEERIMENETYKSIÄ. Epidemiologiset analyysit ovat vaikeita tehdä, koska varhaisen elämän TELOMEERIPITUUS määräytyy perimän mukaan, lähinnä isän puolelta, ja myöhemmin vaikuttuu elämäntyylistä ja ympäristötekijöistä. Lisäksi eräs komplisoiva tekijä on sekin, että YKSILÖLLINEN TELOMEERIPITUUS osittain määräytyy TELOMEERIEN ALKUPITUUDESTA siten, että pitkät TELOMEERIT omaavalla yksilöllä on tietyssä ajanjaksossa kaikkein huomattavin TELOMEERIEN LYHENTYMINEN
En gemensam nämnare för många tillstånd/faktorer som kopplats till kortare telomerer är oxidativ stress, vilket är en känd faktor som kan bidra till ökade telomerförluster. Analyser av epidemiologisk natur är svåra, eftersom vår telomerlängd tidigt i livet till stor del bestäms av arvet (främst från fadern) för att senare påverkas av livsstil och omgivningsfaktorer. Ytterligare en komplicerande faktor är att den individuella telomerförlusten till en del bestäms av ursprungslängden på telomererna, på så sätt att individer med långa telomerer visat den mest uttalade telomerförkortningen under en viss tidsperiod.
(71) Hämmentävä, mutta mielenkiintoinen löytö on sekin, että veren TELOMEERIPITUUS voi olla prognostinen tekijä syövässä. Prognoosi on huonompi niillä, joilla on pitkät telomeeritperifeerisen veren "buffy-coat-soluissa ( jyväisissä valkosoluissa , imusoluissa ja monosyyteissä). Tämä on havaittu rinta-ja munuaissyöpäpotilailta. Molemmsisa ryhmissä oli telomeeripituus riippumaton prognostinen indikaattori. Nämä tiedot osoittavat, että TELOMEERIPITUUS veressä ei ole staattinen, vaan voi vaikuttua tuumoritaudista. Näiden muutosten taustamekanismit ovat monimutkaisia eikä niitä ole vielä selvitetty.
Ett förvånande men intressant fynd är att telomerlängd i blod kan vara en prognostisk faktor vid cancer. Den patientgrupp som hade långa telomerer i buffy coat-celler (granulocyter, lymfocyter och monocyter) från perifert blod befanns nämligen ha sämre prognos än övriga patienter. Detta resultat har erhållits från patienter med såväl bröst- som njurcancer. I båda patientgrupperna var telomerlängd en signifikant oberoende prognostisk indikator [16]. Dessa data tyder ytterligare på att telomerlängden i blod inte är statisk utan kan påverkas, som i detta fall av tumörsjukdom. Mekanismerna bakom dessa förändringar är komplexa och har ännu inte klarlagts.
Kirjoittajien antamat viitteet:
Referenser
1. Shampay J, Szostak JW, Blackburn EH. DNA sequences of telomeres maintained in yeast. Nature. 1984;310(5973):154-7.
2. Szostak JW, Blackburn EH. Cloning yeast telomeres on linear plasmid vectors. Cell. 1982;29(1):245-55.
3. Harley CB, Futcher AB, Greider CW. Telomeres shorten during ageing of human fibroblasts. Nature. 1990;345(6274):458-60.
4. Greider CW, Blackburn EH. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell. 1985;43:405-13.
5. Greider CW, Blackburn EH. A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis. Nature. 1989;337(6205):331-7.
6. de Lange T. Shelterin: the proteincomplex that shapes and safeguards human telomeres. Genes Dev. 2005; 19(18):2100-10.
7. Yamaguchi H, Baerlocher GM, Lansdorp PM, Chanock SJ, Nunez O, Sloand E, et al. Mutations of the human telomerase RNA gene (TERC) in aplastic anemia and myelodysplastic syndrome. Blood. 2003;102(3):916-8.
8. Mitchell JR, Wood E, Collins K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature. 1999;402(6761):551-5.
9. Knight SW, Heiss NS, Vulliamy TJ, Aalfs CM, McMahon C, Richmond P, et al. Unexplained aplastic anaemia, immunodeficiency, and cerebellar hypoplasia (Hoyeraal-Hreidarsson syndrome) due to mutations in the dyskeratosis congenita gene, DKC1. Br J Haematol. 1999;107(2):335-9.
10. Tsakiri KD, Cronkhite JT, Kuan PJ, Xing C, Raghu G, Weissler JC, et al. Adult-onset pulmonary fibrosis caused by mutations in telomerase. Proc Natl Acad Sci U S A. 2007;104(18):7552-7.
11. Mehle C, Ljungberg B, Roos G. Telomere shortening in renal cell carcinoma. Cancer Res. 1994;54(1):236-41.
12. Counter CM, Hirte HW, Bacchetti S, Harley CB. Telomerase activity in human ovarian carcinoma. Proc Natl Acad Sci U S A. 1994;91(8):2900-4.
13. Nilsson P, Mehle C, Remes K, Roos G. Telomerase activity in vivo in human malignant hematopoietic cells. Oncogene. 1994;9(10):3043-8.
14. Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL, et al. Specific association of human telomerase activity with immortal cells and cancer. Science. 1994;266:2011-5.
15. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100(1):57-70.
16. Svenson U, Roos G. Telomere length as a biological marker in malignancy. Biochim Biophys Acta. 2009;1792(4):317-23.
17. Harley CB. Telomerase and cancer therapeutics. Nat Rev Cancer. 2008;8(3);167-79.
KUVAT
Figur 1. Fluorescent in situ-hybridisering (FISH) av normala celler i metafas och interfas. Signalstyrkan är proportionell mot telomerlängden. FISH-preparation: Sofie Degerman, institutionen för medicinsk biovetenskap, Umeå universitet, och Katarina Nordfjäll, ST-läkare, Östersunds sjukhus.
Figur 2. Normala celler förlorar sina telomerer, och maligna, telomeraspositiva celler behåller sina telomerer för varje celldelning. Utan telomeras förkortas kromosomen varje gång cellen delar sig. Slutligen bryts den ner. Telomeras förlänger telomeren och säkerställer därmed att kromosomen inte blir kortare. Grafik: Annika Röhl/Nobelkommittén för fysiologi eller medicin
http://www.lakartidningen.se/store/images/9/9508/large/Fig2ny.jpg
{kind=link}
Figur 3. Principskiss av hur telomeras förlänger telomerer. Angiven DNA-sekvens för telomeren avser Tetrahymena. Enzymet telomeras arbetar på kromosomens ände; det består av ett protein och en RNA-sekvens. RNA-delen är mall när telomer-DNA bildas i änden på kromosomen. Grafik: Annika Röhl/Nobelkommittén för fysiologi eller medicin
http://www.lakartidningen.se/store/images/9/9509/large/Fig3ny.jpg
{kind=link}
Telomeereistä on kiinnostusta ihan Nobel-tasolla
LÄHDE: KLINIK OCH VETENSKAP
Nobelpriset om telomerers skyddsfunktion kan ge stor klinisk nytta
2009 års Nobelpris i fysiologi eller medicin för upptäckten av hur kromosomerna skyddas av telomerer och enzymet telomeras belönar fynd som fått stort genomslag inom skilda biomedicinska forskningsfält. Flera forskningsmål är i sikte, inte bara vad gäller nya cancerterapier.
http://mail.google.com/mail/?hl=sv&tab=wm#inbox/1257a550f23fc9f3
Toistan edellisen kirjoitukseni yhden lähteen:
I6.)Inositolifisfosfaattisignalointi säätelee telomeerien pituutta.
York SJ, Armbruster BN et al. Inositol diphosphate signaling regulates telomere length. J Biol Chem. 2005 Feb 11;280(6):4264-9. Epub 2004 Nov 23.PMID: 15561716
PLC-entsyymistä riippuva inositolifosfaattisignalointitie saa aikaan erilaisia välittäviä signaaleita (messengers), joita muodostuu IP3:sta käsin ja ne kontrolloivat geeniexpressiota ja mRNA:n -kuljetusta ulos tumasta.
PLC =phospholipase C
IP3 on tässä inositol 1,4,5-trisphosphate.
Tässä artikkelissa tutkijat raportoivat telomeerien pituuden säätelyä PP-IP4 molekyylillä, jota tuottaa KCS1 geenituote.
PP-IP4 = diphosphorylinositol tetrakisphosphate.
Jos PP-IP4 tuottuminen poistetaan, telomeerit alkavat muodostua pitemmiksi.
Jos PP-IP4 molekyyliä tuottuu ylimäärin, telomeerit lyhenevät.
Tämä vaikutus vaatii Tel1 läsnäoloa( hiivalla tämä vastaa ihmisen ATM (= protein mutated in the human disease ataxia telangiectasia).
Nämä tiedot osoittavat, että on in vivo olemassa säätelylinkki inositolipolyfosfaattisignaloinnin ja checkpoint kinaasien perheen kesken ja samalla he kuvaavat kolmannen nukleaarisen prosessin, jota moduloi PLC aktivaatio.
KOMMENTTI:
Koska inositoli/fytiini) aineenvaihdunta vaikuttaa telomeeriasiaan, suomennan vähitellen koko Nobel-palkinnon saaneen teleomeeria koskevan aiheiston, vaikka siinä ei mainita sanaa inositoli tai fytiini tai mitään koko PI/IP-alueesta. Sitä paitsi minun täytyy suomentaa se sitäkin varten, että saan paremmin käsitettyä sen kohdan mitä IP/PI modifioi/ moduloi.Onhan IP ravintotekijä myös.
Nobelpriset om telomerers skyddsfunktion kan ge stor klinisk nytta
2009 års Nobelpris i fysiologi eller medicin för upptäckten av hur kromosomerna skyddas av telomerer och enzymet telomeras belönar fynd som fått stort genomslag inom skilda biomedicinska forskningsfält. Flera forskningsmål är i sikte, inte bara vad gäller nya cancerterapier.
http://mail.google.com/mail/?hl=sv&tab=wm#inbox/1257a550f23fc9f3
Toistan edellisen kirjoitukseni yhden lähteen:
I6.)Inositolifisfosfaattisignalointi säätelee telomeerien pituutta.
York SJ, Armbruster BN et al. Inositol diphosphate signaling regulates telomere length. J Biol Chem. 2005 Feb 11;280(6):4264-9. Epub 2004 Nov 23.PMID: 15561716
PLC-entsyymistä riippuva inositolifosfaattisignalointitie saa aikaan erilaisia välittäviä signaaleita (messengers), joita muodostuu IP3:sta käsin ja ne kontrolloivat geeniexpressiota ja mRNA:n -kuljetusta ulos tumasta.
PLC =phospholipase C
IP3 on tässä inositol 1,4,5-trisphosphate.
Tässä artikkelissa tutkijat raportoivat telomeerien pituuden säätelyä PP-IP4 molekyylillä, jota tuottaa KCS1 geenituote.
PP-IP4 = diphosphorylinositol tetrakisphosphate.
Jos PP-IP4 tuottuminen poistetaan, telomeerit alkavat muodostua pitemmiksi.
Jos PP-IP4 molekyyliä tuottuu ylimäärin, telomeerit lyhenevät.
Tämä vaikutus vaatii Tel1 läsnäoloa( hiivalla tämä vastaa ihmisen ATM (= protein mutated in the human disease ataxia telangiectasia).
Nämä tiedot osoittavat, että on in vivo olemassa säätelylinkki inositolipolyfosfaattisignaloinnin ja checkpoint kinaasien perheen kesken ja samalla he kuvaavat kolmannen nukleaarisen prosessin, jota moduloi PLC aktivaatio.
KOMMENTTI:
Koska inositoli/fytiini) aineenvaihdunta vaikuttaa telomeeriasiaan, suomennan vähitellen koko Nobel-palkinnon saaneen teleomeeria koskevan aiheiston, vaikka siinä ei mainita sanaa inositoli tai fytiini tai mitään koko PI/IP-alueesta. Sitä paitsi minun täytyy suomentaa se sitäkin varten, että saan paremmin käsitettyä sen kohdan mitä IP/PI modifioi/ moduloi.Onhan IP ravintotekijä myös.
onsdag 9 december 2009
IP4-PP uutta PubMed lähteestä
Hakusana IP4-PP antaa 17 tulosta PubMed hakulaitteella:
Onnebo SM, Saiardi A. Inositol pyrophosphates modulate hydrogen peroxide signalling.
Biochem J. 2009 Sep 14;423(1):109-18.PMID: 19614566
Inositolipyrofosfaateilla on monia solutoimintoja, mutta erityiset tiet ja downstream-kohteet ovat niukasti luonnehdittuja. Tämä tutkimus on tehty hiivasolussa. Tutkittiin inositolipyrofosfaattien osuutta soluvasteessa, joka muodostuu vapaitten radikaalien (ROS) vaurioitettua solua.
IP6K = inositol hexakisphosphate kinase ; lisää seitsemännen fosfaatin
IP7 = diphosphoinositol pentakisphosphate, IP5-PP
IP8=bisdiphosphoinositol tetrakisphosphate , IP4-(PP)2
PP-IP5K, IP7K, IP7 kinase; lisää kahdeksannen fosfaatin
Jos vip1 puuttuu hiivasolusta , solut akkumuloivat suuret määrät IP7-muotoa , mutta IP8 ei muodostu, joten tämä entsyymi edustaa fysiologista IP7-kinaasia.
Vasta IP8- muoto ( tuplasti pyrofosforyloitunut muoto IP8 , [(PP)2-IP4]), välittää vasteen vahingollisen vetyperoksidin muodostumiselle ja saa solut resistenteiksi H2O2:n aiheuttamalle tuholle.
Kehossa kuluu nopeasti inositolipyrofosfaattia, jos on altistusta vetyperoksidille. ROS-signaali välittyy evolutionaalisesti konservatiivisen entsyymiluokan suoralla modifikaatiolla (siinä on cysteiini).
Structural analysis and detection of biological inositol pyrophosphates reveal that the family of VIP/diphosphoinositol pentakisphosphate kinases are 1/3-kinases.
J Biol Chem. 2009 Jan 16;284(3):1863-72. Epub 2008 Nov 3. PMID: 18981179
1/3,5-(PP)2-IP4 on isomeerinen rakenne bis-difosfoinositoli-tetrakisfosfaattia (IP8) , jota syntetisoituu niin hiivassa kuin ihmisessä IP6K entsyymin ja VIP/PP-IP5K entsyymien yhteistyönä. Löydettiin fylogeneettistä vaihtelevuutta inositolipyrofosfaattien taksonomiassa.
Sellaisessa lajissa kuin Dictyostelium (eräs limasienilaji) esiintyi lähinnä muotoa bis-diphosphoinositol tetrakisphosphate ins 5,6-(PP)2-IP4.Tutkijat koettivat löytää rakenteen ja funktion suhteita, jotka kontrolloivat inositolipyrofosfaattien synteesin ja biologisten toimintojen spesifisyyttä.
Fridy PC, Otto JC, Dollins DE, York JD.
Cloning and characterization of two human VIP1-like inositol hexakisphosphate and diphosphoinositol pentakisphosphate kinases.
J Biol Chem. 2007 Oct 19;282(42):30754-62. Epub 2007 Aug 9.PMID: 17690096
EUKARYOOTEILLA on lukuisia inositolifosfaatteja (IP) ja difosfoinositolifosfaatteja (PP-IPs), joita myös kutsutaan inositolipyrofosfaateiksi ja ne vaikuttavat kemiallisina koodeina, jotka ovat tärkeitä solunsisäisissä signaaliteissä. IP- ja PP-IP-molekyylijoukkoa tuottaa useat evolutionaalisesti konservoidut IP-kinaasit.
Tässä tutkijat kertovat, miten he saivat luonnehdittua IP6-kinaasin (IP6K) ja IP7-kinaasin (IP7K= diphosphoinositol pentakisphosphate eli PP-IP5 kinase), jotka olivat hiivassa sen Vip1- entsyymin kaltaisia ja ne identifioitiin IP6/IP7 kinaasina. Koeputkessa VIP1 osoittaa IP6kinaasi - ja IP7-kinaasiaktiivisuuta ja saa aikaan IP7 synteesin. VIP1 geenin ilmenemä ihmissolussa tuotti korkeat pitoisuudet IP7 , josta taas johtui dramaattinen lisääntymä IP8-muodossa (= bisdiphosphoinositol tetrakisphosphate (PP2-IP4 or IP8).
Missä ihmiskehon kohdassa ilmenee VIP1 geeniä? Sitä on monissa eri kudoksissa ja erityisen rikkaana tahdonalaisessa lihaksessa, sydämessä ja aivoissa. Jos VIP1 merkattiin ja katsottiin, missä sitä on, niin sitä löydettiin solun sytoplasman non-membraanisessa tilassa, kohdissa missä ei ole kalvorakenteita.
Tutkijat luonnehtivat myös ihmisen ja hiiren VIP2-entsyymin, joka on toisen geenin tuote, mutta 90 %:sti samanlainen kuin VIP1-entsyymi kinaasidomaaniltaan ja se osoitti myös sekä IP6K ja IP7K aktiivisuutta. Tutkijat päättelivät, että ihmisen VIP1 ja VIP2 toimivat IP6- ja IP7-kinaaseina, jotka vaikuttavat yhteistyössä IP6K/Kcs1-luokan kinaasien kanssa konvertoimassa IP6-muotoa nisäkässoluissa IP8-muotoon. Tämän prosessin on todettu tapahtuvan vasteena erilaisille stimuluksille ja signalointitapahtumille.
4. Hiivasolussa on tehty (2005) tutkimus Ins-(1,4,5)-P3 inositolitrifosfaatin aloittaman (uuden) inositolipolyfostaattitien molekulaariseksi määrittämiseksi.
LÄHDE:
Seeds AM, Bastidas RJ, York JD. Molecular definition of a novel inositol polyphosphate (IPx) metabolic pathway initiated by inositol 1,4,5-trisphosphate 3-kinase activity in Saccharomyces cerevisiae. J Biol Chem. 2005 Jul 29;280(30):27654-61. Epub 2005 Jun 8.PMID: 15944147
Inositolipolyfosfaatteja (IPs) ja inositolipyrofosfaatteja (PP-IPs syntetisoituu inositolitrifosfaatista käsin, mutta on ajateltu että IP3 täytyy olla silloin muodossa inositoli-1,4,5-trifosfaatti ( eli I(1,4,5)P3).
Tähän alkumuokkaukseen tarvitaan Ipk2 kinaasiaktiivisuudet: 6-/3-/5-kinase.
Tämä kinaasi on tunnettu myös nimellä Arg82 tai sitten multikinaasina eli nimellä inositol polyphosphate multikinase..
Tässä työssään tutkijat katsoivat IP-aineenvaihdunnassa esiintyviä eroja mainittujen 6- ja 3- kinaasien kesken (distinct roles for I(1,4,5)P3 6- versus 3-kinase activities in IP metabolism and cellular functions reported for Ipk2).
3-kinaasi aloitti uuden synteesitien , josta tuli yli 11 kpl. IPs ja PP-IPs.
Primääri synteesitie oli
I(1,4,5)P3
I(1,3,4,5)P4
I(1,2,3,4,5)P5
PP-IP4 ,
PP2-IP3 ja vaati Kcs1 (tai mahdollisesti Ipk2).
Tutkijat tekivät päättelyn myös uuden IP3-kinaasin olemassaolosta, jolla on merkitystä nukleosytoplasmisissa prosesseissa ja se vaikuttaa IPs ja PP-IPs molekyylimäärien keskinäisiin kompensaatioihin ( toisiaan korvaavaisuuteen).
Zanelli SA, Spandou E et al. Hypoxia modifies nuclear calcium uptake pathways in the cerebral cortex of the guinea-pig fetus. Neuroscience. 2005;130(4):949-55.PMID: 15652992
York SJ, Armbruster BN et al. Inositol diphosphate signaling regulates telomere length. J Biol Chem. 2005 Feb 11;280(6):4264-9. Epub 2004 Nov 23.PMID: 15561716
PLC-entsyymistä riippuva inositolifosfaattisignalointitie saa aikaan erilaisia välittäviä signaaleita (messengers), joita muodostuu IP3:sta käsin ja ne kontrolloivat geeniexpressiota ja mRNA:n -kuljetusta ulos tumasta.
PLC =phospholipase C
IP3 on tässä inositol 1,4,5-trisphosphate.
Tässä artikkelissa tutkijat raportoivat telomeerien pituuden säätelyä PP-IP4 molekyylillä, jota tuottaa KCS1 geenituote.
PP-IP4 = diphosphorylinositol tetrakisphosphate.
Jos PP-IP4 tuottuminen katoaa, telomeerit alkavat muodostua pitemmiksi.
Jos PP-IP4 molekyyliä tuottuu ylimäärin, telomeerit lyhenevät.
Tämä vaikutus vaatii Tel1 läsnäoloa( hiivalla tämä vastaa ihmisen ATM (= protein mutated in the human disease ataxia telangiectasia).
Nämä tiedot osoittavat, että on in vivo olemassa säätelylinkki inositolipolyfosfaattisignaloinnin ja checkpoint kinaasien perheen kesken ja samalla kuvaavat kolmannen nukleaarisen prosessin, jota moduloi PLC aktivaatio.
7. Banaanikärpäsessä tehty tutkimus inositolipolyfosfosfaattien synteesistä molekyylitasolla.
Seeds AM, Sandquist JC et al. A molecular basis for inositol polyphosphate synthesis in Drosophila melanogaster. J Biol Chem. 2004 Nov 5;279(45):47222-32. Epub 2004 Aug 20.PMID: 15322119
8. Tutkimusta difosfoinositolipentakisfosfaattikinaasista Se syntetisoi inositolipyrofosfaattia, bis(difosfo) inositolitetrakisfosfaattia, IP4-PP.
Huang CF, Voglmaier SM et al. Identification and purification of diphosphoinositol pentakisphosphate kinase, which synthesizes the inositol pyrophosphate bis(diphospho)inositol tetrakisphosphate. Biochemistry. 1998 Oct 20;37(42):14998-5004.PMID: 9778378
9. Motorisen neutronin taudissa esiintyvä hajuradan häiriö.
Hawkes CH, Shephard BC et al. Olfactory disorder in motor neuron disease. Exp Neurol. 1998 Apr;150(2):248-53.PMID: 9527894
10. alfa1-adrenoreseptoristimulaatio estää osittain ATP-herkkää kaliumvirtaa koe-eläimen ventrikulaarisessa solussa.
Takizawa T, Hara Y et al. alpha 1-Adrenoceptor stimulation partially inhibits ATP-sensitive K+ current in guinea pig ventricular cells: attenuation of the action potential shortening induced by hypoxia and K+ channel openers. J Cardiovasc Pharmacol. 1996 Dec;28(6):799-808.PMID: 8961078
11. D-myo-inositolijohdannaisia pienissä unilamellaarisissa liposomeissa.
Brailoiu E, Beschea C et al.
TLC characterization of small unilamellar liposomes containing D-myo-inositol derivatives.
Biomed Chromatogr. 1996 Sep-Oct;10(5):233-6.PMID: 8879530
12.IP6-kinaasi on ATP-syntaasi: difosfoinositolipentakisfosfaatti korkea-energisen fosfaatin luovuttajana
Voglmaier SM, Bembenek ME et al.
Purified inositol hexakisphosphate kinase is an ATP synthase: diphosphoinositol pentakisphosphate as a high-energy phosphate donor.
Proc Natl Acad Sci U S A. 1996 Apr 30;93(9):4305-10.PMID: 8633060
15. Sydäntä suojaava vaikutus korreloi lisääntyneeseen ekto-5´nukleotidaasiaktiivisuuteen.
Minamino T, Kitakaze M et al. Cardioprotection due to preconditioning correlates with increased ecto-5'-nucleotidase activity. Am J Physiol. 1996 Jan;270(1 Pt 2):H238-44.PMID: 8769757
14. D-myo-inositolijohdannaisten luonnehdinta liposomissa.
Brailoiu E, Huhurez G et al.
TLC characterization of liposomes containing D-myo-inositol derivatives.
Biomed Chromatogr. 1995 Jul-Aug;9(4):175-8.PMID: 8520206
15.Golgi co-atomit sitovat ja muodostavat kaliumselektiivisiä kanavia, joita ohjaa inositolipolyfosfaatit.
Fleischer B, Xie J, Mayrleitner M et al. Golgi coatomer binds, and forms K(+)-selective channels gated by, inositol polyphosphates. J Biol Chem. 1994 Jul 8;269(27):17826-32.PMID: 8027036
16. Koe-eläimen sydämen IP3-kinaasiaktiivisuus
Kawaguchi H, Iizuka K et al. Inositol trisphosphate kinase activity in hypertrophied rat heart.
Biochem Med Metab Biol. 1990 Aug;44(1):42-50.PMID: 2167704
17. Ins-1,3,4-P3 muunnellee kalvopotentiaalia .
Tertoolen LG, Tilly BC et al.
Electrophysiological responses to bradykinin and microinjected inositol polyphosphates in neuroblastoma cells. Possible role of inositol 1,3,4-trisphosphate in altering membrane potential.
FEBS Lett. 1987 Apr 20;214(2):365-9.PMID: 3494634
1. Inositolipyrofosfaatit moduloivat H2O2 (vetyperoksidi) signalointia
LÄHDE:Onnebo SM, Saiardi A. Inositol pyrophosphates modulate hydrogen peroxide signalling.
Biochem J. 2009 Sep 14;423(1):109-18.PMID: 19614566
Inositolipyrofosfaateilla on monia solutoimintoja, mutta erityiset tiet ja downstream-kohteet ovat niukasti luonnehdittuja. Tämä tutkimus on tehty hiivasolussa. Tutkittiin inositolipyrofosfaattien osuutta soluvasteessa, joka muodostuu vapaitten radikaalien (ROS) vaurioitettua solua.
IP6K = inositol hexakisphosphate kinase ; lisää seitsemännen fosfaatin
IP7 = diphosphoinositol pentakisphosphate, IP5-PP
IP8=bisdiphosphoinositol tetrakisphosphate , IP4-(PP)2
PP-IP5K, IP7K, IP7 kinase; lisää kahdeksannen fosfaatin
Jos vip1 puuttuu hiivasolusta , solut akkumuloivat suuret määrät IP7-muotoa , mutta IP8 ei muodostu, joten tämä entsyymi edustaa fysiologista IP7-kinaasia.
Vasta IP8- muoto ( tuplasti pyrofosforyloitunut muoto IP8 , [(PP)2-IP4]), välittää vasteen vahingollisen vetyperoksidin muodostumiselle ja saa solut resistenteiksi H2O2:n aiheuttamalle tuholle.
Kehossa kuluu nopeasti inositolipyrofosfaattia, jos on altistusta vetyperoksidille. ROS-signaali välittyy evolutionaalisesti konservatiivisen entsyymiluokan suoralla modifikaatiolla (siinä on cysteiini).
2. Biologisten inositolipyrofosfaattien (IPx-PP) rakenteen analyysit ja havaitsemiset paljastavat, että VIP/difosfoinositolipentakisfosfaatti -kinaasit ovat 1/3-kinaaseja
LÄHDE: Lin H, Fridy PC, Ribeiro AA et al.Structural analysis and detection of biological inositol pyrophosphates reveal that the family of VIP/diphosphoinositol pentakisphosphate kinases are 1/3-kinases.
J Biol Chem. 2009 Jan 16;284(3):1863-72. Epub 2008 Nov 3. PMID: 18981179
1/3,5-(PP)2-IP4 on isomeerinen rakenne bis-difosfoinositoli-tetrakisfosfaattia (IP8) , jota syntetisoituu niin hiivassa kuin ihmisessä IP6K entsyymin ja VIP/PP-IP5K entsyymien yhteistyönä. Löydettiin fylogeneettistä vaihtelevuutta inositolipyrofosfaattien taksonomiassa.
Sellaisessa lajissa kuin Dictyostelium (eräs limasienilaji) esiintyi lähinnä muotoa bis-diphosphoinositol tetrakisphosphate ins 5,6-(PP)2-IP4.Tutkijat koettivat löytää rakenteen ja funktion suhteita, jotka kontrolloivat inositolipyrofosfaattien synteesin ja biologisten toimintojen spesifisyyttä.
3. Artikkeli kertoo ihmisten kahden VIP1-kaltaisen kinaasin, IP6-kinaasin (IP6K) ja difosfoinositoli -IP5-kinaasin (IP7K), kloonaamisesta ja luonnehtimisesta.
LÄHDE:Fridy PC, Otto JC, Dollins DE, York JD.
Cloning and characterization of two human VIP1-like inositol hexakisphosphate and diphosphoinositol pentakisphosphate kinases.
J Biol Chem. 2007 Oct 19;282(42):30754-62. Epub 2007 Aug 9.PMID: 17690096
EUKARYOOTEILLA on lukuisia inositolifosfaatteja (IP) ja difosfoinositolifosfaatteja (PP-IPs), joita myös kutsutaan inositolipyrofosfaateiksi ja ne vaikuttavat kemiallisina koodeina, jotka ovat tärkeitä solunsisäisissä signaaliteissä. IP- ja PP-IP-molekyylijoukkoa tuottaa useat evolutionaalisesti konservoidut IP-kinaasit.
Tässä tutkijat kertovat, miten he saivat luonnehdittua IP6-kinaasin (IP6K) ja IP7-kinaasin (IP7K= diphosphoinositol pentakisphosphate eli PP-IP5 kinase), jotka olivat hiivassa sen Vip1- entsyymin kaltaisia ja ne identifioitiin IP6/IP7 kinaasina. Koeputkessa VIP1 osoittaa IP6kinaasi - ja IP7-kinaasiaktiivisuuta ja saa aikaan IP7 synteesin. VIP1 geenin ilmenemä ihmissolussa tuotti korkeat pitoisuudet IP7 , josta taas johtui dramaattinen lisääntymä IP8-muodossa (= bisdiphosphoinositol tetrakisphosphate (PP2-IP4 or IP8).
Missä ihmiskehon kohdassa ilmenee VIP1 geeniä? Sitä on monissa eri kudoksissa ja erityisen rikkaana tahdonalaisessa lihaksessa, sydämessä ja aivoissa. Jos VIP1 merkattiin ja katsottiin, missä sitä on, niin sitä löydettiin solun sytoplasman non-membraanisessa tilassa, kohdissa missä ei ole kalvorakenteita.
Tutkijat luonnehtivat myös ihmisen ja hiiren VIP2-entsyymin, joka on toisen geenin tuote, mutta 90 %:sti samanlainen kuin VIP1-entsyymi kinaasidomaaniltaan ja se osoitti myös sekä IP6K ja IP7K aktiivisuutta. Tutkijat päättelivät, että ihmisen VIP1 ja VIP2 toimivat IP6- ja IP7-kinaaseina, jotka vaikuttavat yhteistyössä IP6K/Kcs1-luokan kinaasien kanssa konvertoimassa IP6-muotoa nisäkässoluissa IP8-muotoon. Tämän prosessin on todettu tapahtuvan vasteena erilaisille stimuluksille ja signalointitapahtumille.
4. Hiivasolussa on tehty (2005) tutkimus Ins-(1,4,5)-P3 inositolitrifosfaatin aloittaman (uuden) inositolipolyfostaattitien molekulaariseksi määrittämiseksi.
LÄHDE:
Seeds AM, Bastidas RJ, York JD. Molecular definition of a novel inositol polyphosphate (IPx) metabolic pathway initiated by inositol 1,4,5-trisphosphate 3-kinase activity in Saccharomyces cerevisiae. J Biol Chem. 2005 Jul 29;280(30):27654-61. Epub 2005 Jun 8.PMID: 15944147
Inositolipolyfosfaatteja (IPs) ja inositolipyrofosfaatteja (PP-IPs syntetisoituu inositolitrifosfaatista käsin, mutta on ajateltu että IP3 täytyy olla silloin muodossa inositoli-1,4,5-trifosfaatti ( eli I(1,4,5)P3).
Tähän alkumuokkaukseen tarvitaan Ipk2 kinaasiaktiivisuudet: 6-/3-/5-kinase.
Tämä kinaasi on tunnettu myös nimellä Arg82 tai sitten multikinaasina eli nimellä inositol polyphosphate multikinase..
Tässä työssään tutkijat katsoivat IP-aineenvaihdunnassa esiintyviä eroja mainittujen 6- ja 3- kinaasien kesken (distinct roles for I(1,4,5)P3 6- versus 3-kinase activities in IP metabolism and cellular functions reported for Ipk2).
3-kinaasi aloitti uuden synteesitien , josta tuli yli 11 kpl. IPs ja PP-IPs.
Primääri synteesitie oli
I(1,4,5)P3
I(1,3,4,5)P4
I(1,2,3,4,5)P5
PP-IP4 ,
PP2-IP3 ja vaati Kcs1 (tai mahdollisesti Ipk2).
Tutkijat tekivät päättelyn myös uuden IP3-kinaasin olemassaolosta, jolla on merkitystä nukleosytoplasmisissa prosesseissa ja se vaikuttaa IPs ja PP-IPs molekyylimäärien keskinäisiin kompensaatioihin ( toisiaan korvaavaisuuteen).
5. Koe-eläimen sikiön aivokuoren nukleaarista kalsiumin ottoa modifioi hypoksia, hapenpuute.
LÄHDE:Zanelli SA, Spandou E et al. Hypoxia modifies nuclear calcium uptake pathways in the cerebral cortex of the guinea-pig fetus. Neuroscience. 2005;130(4):949-55.PMID: 15652992
6. Inositolifisfosfaattisignalointi säätelee telomeerien pituutta.
LÄHDE:York SJ, Armbruster BN et al. Inositol diphosphate signaling regulates telomere length. J Biol Chem. 2005 Feb 11;280(6):4264-9. Epub 2004 Nov 23.PMID: 15561716
PLC-entsyymistä riippuva inositolifosfaattisignalointitie saa aikaan erilaisia välittäviä signaaleita (messengers), joita muodostuu IP3:sta käsin ja ne kontrolloivat geeniexpressiota ja mRNA:n -kuljetusta ulos tumasta.
PLC =phospholipase C
IP3 on tässä inositol 1,4,5-trisphosphate.
Tässä artikkelissa tutkijat raportoivat telomeerien pituuden säätelyä PP-IP4 molekyylillä, jota tuottaa KCS1 geenituote.
PP-IP4 = diphosphorylinositol tetrakisphosphate.
Jos PP-IP4 tuottuminen katoaa, telomeerit alkavat muodostua pitemmiksi.
Jos PP-IP4 molekyyliä tuottuu ylimäärin, telomeerit lyhenevät.
Tämä vaikutus vaatii Tel1 läsnäoloa( hiivalla tämä vastaa ihmisen ATM (= protein mutated in the human disease ataxia telangiectasia).
Nämä tiedot osoittavat, että on in vivo olemassa säätelylinkki inositolipolyfosfaattisignaloinnin ja checkpoint kinaasien perheen kesken ja samalla kuvaavat kolmannen nukleaarisen prosessin, jota moduloi PLC aktivaatio.
7. Banaanikärpäsessä tehty tutkimus inositolipolyfosfosfaattien synteesistä molekyylitasolla.
Seeds AM, Sandquist JC et al. A molecular basis for inositol polyphosphate synthesis in Drosophila melanogaster. J Biol Chem. 2004 Nov 5;279(45):47222-32. Epub 2004 Aug 20.PMID: 15322119
8. Tutkimusta difosfoinositolipentakisfosfaattikinaasista Se syntetisoi inositolipyrofosfaattia, bis(difosfo) inositolitetrakisfosfaattia, IP4-PP.
Huang CF, Voglmaier SM et al. Identification and purification of diphosphoinositol pentakisphosphate kinase, which synthesizes the inositol pyrophosphate bis(diphospho)inositol tetrakisphosphate. Biochemistry. 1998 Oct 20;37(42):14998-5004.PMID: 9778378
9. Motorisen neutronin taudissa esiintyvä hajuradan häiriö.
Hawkes CH, Shephard BC et al. Olfactory disorder in motor neuron disease. Exp Neurol. 1998 Apr;150(2):248-53.PMID: 9527894
10. alfa1-adrenoreseptoristimulaatio estää osittain ATP-herkkää kaliumvirtaa koe-eläimen ventrikulaarisessa solussa.
Takizawa T, Hara Y et al. alpha 1-Adrenoceptor stimulation partially inhibits ATP-sensitive K+ current in guinea pig ventricular cells: attenuation of the action potential shortening induced by hypoxia and K+ channel openers. J Cardiovasc Pharmacol. 1996 Dec;28(6):799-808.PMID: 8961078
11. D-myo-inositolijohdannaisia pienissä unilamellaarisissa liposomeissa.
Brailoiu E, Beschea C et al.
TLC characterization of small unilamellar liposomes containing D-myo-inositol derivatives.
Biomed Chromatogr. 1996 Sep-Oct;10(5):233-6.PMID: 8879530
12.IP6-kinaasi on ATP-syntaasi: difosfoinositolipentakisfosfaatti korkea-energisen fosfaatin luovuttajana
Voglmaier SM, Bembenek ME et al.
Purified inositol hexakisphosphate kinase is an ATP synthase: diphosphoinositol pentakisphosphate as a high-energy phosphate donor.
Proc Natl Acad Sci U S A. 1996 Apr 30;93(9):4305-10.PMID: 8633060
15. Sydäntä suojaava vaikutus korreloi lisääntyneeseen ekto-5´nukleotidaasiaktiivisuuteen.
Minamino T, Kitakaze M et al. Cardioprotection due to preconditioning correlates with increased ecto-5'-nucleotidase activity. Am J Physiol. 1996 Jan;270(1 Pt 2):H238-44.PMID: 8769757
14. D-myo-inositolijohdannaisten luonnehdinta liposomissa.
Brailoiu E, Huhurez G et al.
TLC characterization of liposomes containing D-myo-inositol derivatives.
Biomed Chromatogr. 1995 Jul-Aug;9(4):175-8.PMID: 8520206
15.Golgi co-atomit sitovat ja muodostavat kaliumselektiivisiä kanavia, joita ohjaa inositolipolyfosfaatit.
Fleischer B, Xie J, Mayrleitner M et al. Golgi coatomer binds, and forms K(+)-selective channels gated by, inositol polyphosphates. J Biol Chem. 1994 Jul 8;269(27):17826-32.PMID: 8027036
16. Koe-eläimen sydämen IP3-kinaasiaktiivisuus
Kawaguchi H, Iizuka K et al. Inositol trisphosphate kinase activity in hypertrophied rat heart.
Biochem Med Metab Biol. 1990 Aug;44(1):42-50.PMID: 2167704
17. Ins-1,3,4-P3 muunnellee kalvopotentiaalia .
Tertoolen LG, Tilly BC et al.
Electrophysiological responses to bradykinin and microinjected inositol polyphosphates in neuroblastoma cells. Possible role of inositol 1,3,4-trisphosphate in altering membrane potential.
FEBS Lett. 1987 Apr 20;214(2):365-9.PMID: 3494634
Prenumerera på:
Inlägg (Atom)