
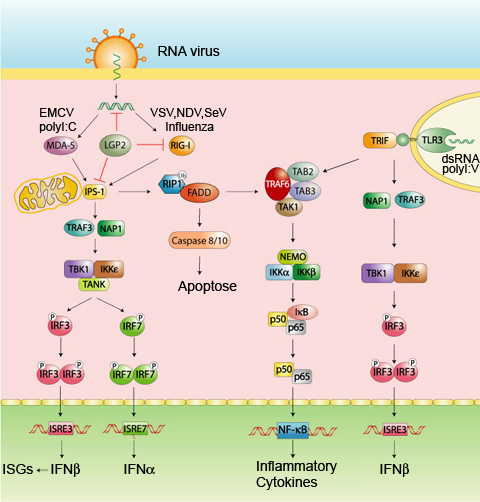
PI3K aktivoituu influenssaviruksen vRNA materiaalista patogeenin rakenteen tunnistavanRNA helikaasi rakenteisen Rig-1 reseptorin kautta mikä edistää tehokkaan I tyypin interferonin muodostusta.
LÄHDE:
Cell Microbiol. 2011 Sep 8. doi: 10.1111/j.1462-5822.2011.01680.x. [Epub ahead of print] Phosphatidylinositol-3-kinase (PI3K) is activated by influenza virus vRNA via the pathogen pattern receptor Rig-I to promote efficient type I interferon production.
Source
Institute of Molecular Virology (IMV), ZMBE, Von Esmarch-Str. 56, D-48149 Muenster, Germany.
Abstract, Tiivistelmä
The phosphatidylinositol-3-kinase (PI3K) was identified to be activated upon influenza A virus infection.
On tunnistettu PI3K entsyymin aktivoituvan A-influenssavirusinfektion aikana.
An early and transient induction of PI3K signaling is caused by viral attachment to cells and promotes virus entry.
Varhaisen ja ohimenevän PI3K-entsyymin aktivoitumisen aiheuttaa viruksen liittyminen soluun ja tässä PI3K- aktivaatio edistää viruksen sisäänpääsyä.
In later phases of infection the kinase is activated by the viral NS1 protein to prevent premature apoptosis.
Infektion myöhemmässä vaiheessa viruksen NS1-proteiini aktivoi PI3K enstyymiä estääkseen isäntäsolun ennenaikaisen apoptoosin.
Besides these virus supporting functions, it was suggested that PI3K signaling is involved in dsRNA and IAV induced antiviral responses by enhancing the activity of interferon regulatory factor-3 (IRF-3).
Näitten virukselle edullisten funktioitten lisäksi PI3K-signalointi osallistuu viruksen dsRNA muodon ja A-influenssaviruksen indusoimiin antivirusvasteisiin lisäämällä interferonia säätelevän tekijä 3:n (IRF-3) aktiivisuutta.
However, molecular mechanisms of activation remained obscure.
Kuitenkin taustalla oleva molekulaarinen mekanismi on pysynyt hämärän peitossa.
Here we show that accumulation of vRNA in cells infected with influenza A or B viruses results in PI3K activation.
Tässä tutkimuksessa tiedemiehet osoittivat, että influenssa A tai B virusten infektoimissa soluissa kertyvä virusRNA johtaa PI3K-entsyymin aktivoitumiseen.
Furthermore, expression of the RNA receptors Rig-I and MDA5 was increased upon stimulation with virion extracted vRNA or IAV infection.
Lisäksi virionista uutettu vRNA tai influenssavirus-infektio sinänä stimuloivat lisääntynyttä RNA-reseptorien Rig-1 ja MDA5 ilmenemää.
Using siRNA approaches, Rig-I was identified as pathogen receptor necessary for influenza virus vRNA sensing and subsequent PI3K activation in a TRIM25 and MAVS signaling dependent manner.
Kun tehtiin tutkimuksia siRNA:lla, todettiin Rig-1 patogeenireseptoriksi, joka oli välttämätön viruksen vRNA muodon tunnistamisessa ja sitä seuraavassa PI3K-aktivaatiossa tavalla, joka riippui TRIM25 ja MAVS signaloinnista.
Rig-I induced PI3K signaling was further shown to be essential for complete IRF-3 activation and consequently induction of the type I interferon response.
Lisäksi osoittautui, että Rig-I:n indusoima PI3kinaasin signalointi oli välttämätön, jotta IRF-3 saattoi aktivoitua täydellisesti ja saada aikaan tyypin I interferonivasteen.
These data identify PI3K as factor that is activated as part of the Rig-I mediated anti-pathogen response to enhance expression of type I interferons.
Näistä tiedoista päätellen PI3K on tekijä, joka aktivoituu osana Rig-1 välitteisestä antipatogeenivasteesta I-tyyppisten interferonien expression lisäämisessä.
Rig-1
http://www.nature.com/ng/journal/v38/n8/images/ng0806-866-F1.jpg
{kind=link}
IRF
{kind=link}
http://www.nature.com/ni/journal/v5/n7/full/ni1087.html